Você também pode gostar
- Resumo 3 - Nutrição MineralDocumento6 páginasResumo 3 - Nutrição MineralLuana LimaAinda não há avaliações
- Micronutrientes: Funções do manganês nas plantasDocumento6 páginasMicronutrientes: Funções do manganês nas plantasMoniqueMacedoAinda não há avaliações
- Nutrição mineral essencial para plantasDocumento49 páginasNutrição mineral essencial para plantasAlinfur100% (1)
- Aposti6 - Nutrição Mineral de PlantasDocumento5 páginasAposti6 - Nutrição Mineral de Plantasluan_aliAinda não há avaliações
- Tabelas Sobre Macro e MicronutrientesDocumento5 páginasTabelas Sobre Macro e MicronutrientesJhonas Aikanã KassupáAinda não há avaliações
- ESTUDO DIRIGIDO - Nutrição MineralDocumento4 páginasESTUDO DIRIGIDO - Nutrição MineralAckça PriscilaAinda não há avaliações
- Nutrição Mineral de PlantasDocumento48 páginasNutrição Mineral de PlantasNívia IvoAinda não há avaliações
- Importância Dos NutirentesDocumento5 páginasImportância Dos NutirentesPriscila JulianiAinda não há avaliações
- Neloyd 1Documento19 páginasNeloyd 1Sulemane IssumalgyAinda não há avaliações
- Elementos benéficos nas plantas: sódio, silício e cobaltoDocumento6 páginasElementos benéficos nas plantas: sódio, silício e cobaltoKelly SousaAinda não há avaliações
- Importância dos nutrientes minerais para a nutrição e produtividade das plantasDocumento12 páginasImportância dos nutrientes minerais para a nutrição e produtividade das plantasBenjamin LawsonAinda não há avaliações
- Minerais Form Adores de RochasDocumento11 páginasMinerais Form Adores de RochasPaulo FreireAinda não há avaliações
- Elementos Benéficos e Tóxicos no SoloDocumento47 páginasElementos Benéficos e Tóxicos no SoloJeferson FerreiraAinda não há avaliações
- Importância dos Nutrientes Minerais para as PlantasDocumento7 páginasImportância dos Nutrientes Minerais para as PlantasSheyenne CarvalhoAinda não há avaliações
- Constance SlidesCarnivalDocumento20 páginasConstance SlidesCarnivalJúlia MariaAinda não há avaliações
- Atividade 1 - Nutrição e FotossinteseDocumento11 páginasAtividade 1 - Nutrição e Fotossintesea097792Ainda não há avaliações
- NUTRIÇÃODocumento6 páginasNUTRIÇÃOAgronomia UnoparAinda não há avaliações
- Adubação FoliarDocumento8 páginasAdubação FoliarFabíola EstrelaAinda não há avaliações
- Nutrição Mineral Das Plantas - Revisão de LiteraturaDocumento10 páginasNutrição Mineral Das Plantas - Revisão de LiteraturaVítor OliveiraAinda não há avaliações
- Fertilidade Do Solo Aula XIIDocumento49 páginasFertilidade Do Solo Aula XIIFernando Eugenio da SilvaAinda não há avaliações
- Nutrição Mineral em PlantasDocumento2 páginasNutrição Mineral em PlantasTathiane Dias84% (25)
- Micronutrientes - FerroDocumento5 páginasMicronutrientes - FerroMoniqueMacedoAinda não há avaliações
- Nutrientes essenciais plantasDocumento11 páginasNutrientes essenciais plantascarllosmagnoAinda não há avaliações
- Fndamentos de ZoologiaDocumento124 páginasFndamentos de ZoologiaAmanda SchmidtAinda não há avaliações
- 2017 - Bila - Elementos Tóxicos As PlantasDocumento45 páginas2017 - Bila - Elementos Tóxicos As PlantasMarciel Souza CarvalhoAinda não há avaliações
- Critérios de EssencialidadeDocumento7 páginasCritérios de EssencialidadeEdson Acácio Muianga MuiangaAinda não há avaliações
- Causas de Alcalinidade Do SoloDocumento6 páginasCausas de Alcalinidade Do SoloNunoClaudinoAinda não há avaliações
- Deficiências de micronutrientes no cafeeiroDocumento6 páginasDeficiências de micronutrientes no cafeeiroEveraldo Borges da CostaAinda não há avaliações
- Micronutrientes - Nutrição Mineral de Plantas - FERNANDES, M. S. - SBCSDocumento27 páginasMicronutrientes - Nutrição Mineral de Plantas - FERNANDES, M. S. - SBCSLeandro Moscoso50% (2)
- Nutrição de plantas e seus nutrientesDocumento3 páginasNutrição de plantas e seus nutrientesJúnior FogaçaAinda não há avaliações
- Nutrição MineralDocumento58 páginasNutrição MineralGlauco Naressi100% (3)
- Micronutrientes em plantasDocumento27 páginasMicronutrientes em plantasCamila FerreiraAinda não há avaliações
- Micronutrientes essenciais: funções, sintomas de deficiência e excessoDocumento19 páginasMicronutrientes essenciais: funções, sintomas de deficiência e excessoGustavo de BarrosAinda não há avaliações
- Os metais no solo: origem, transformações e impactos ambientaisDocumento4 páginasOs metais no solo: origem, transformações e impactos ambientaisAgrovale Projetos agropecuários e ambientaisAinda não há avaliações
- Minerais e NutrientesDocumento4 páginasMinerais e NutrientesCurso SobrancelhaAinda não há avaliações
- Estudo Dirigido AP1 Cultura de Tecidos e Biofábricas BiotecnologiaDocumento25 páginasEstudo Dirigido AP1 Cultura de Tecidos e Biofábricas BiotecnologiaMilena MacielAinda não há avaliações
- Aula Nutrição Mineral - CópiaDocumento69 páginasAula Nutrição Mineral - CópiaGabriel ArensAinda não há avaliações
- Nutrição Mineral de Plantas: FertilidadeDocumento7 páginasNutrição Mineral de Plantas: FertilidadeDon LucasAinda não há avaliações
- Nutrientes No SoloDocumento10 páginasNutrientes No SoloRoberto Rac CorreaAinda não há avaliações
- Cobre - Fisiologia VegetalDocumento21 páginasCobre - Fisiologia VegetalElano CavalcantiAinda não há avaliações
- Nutricao Mineral de PlantasDocumento30 páginasNutricao Mineral de PlantasHugo Henrique100% (1)
- Nutrição mineral essencial para plantas: macro e micronutrientesDocumento32 páginasNutrição mineral essencial para plantas: macro e micronutrientesFlavia Fernandes Ribeiro De MirandaAinda não há avaliações
- NutrientesDocumento22 páginasNutrientescagmbrAinda não há avaliações
- Ferro e ManganesDocumento33 páginasFerro e ManganesDaniel ArrudaAinda não há avaliações
- Fosforo e EnxofreDocumento4 páginasFosforo e EnxofreJosé RanonAinda não há avaliações
- F VegetalDocumento2 páginasF VegetalwillycarlosjoaoraulAinda não há avaliações
- ZincoDocumento8 páginasZincoLeila PereiraAinda não há avaliações
- Fundamentos NutriçãoDocumento64 páginasFundamentos NutriçãoNeder BlancoAinda não há avaliações
- Aula 1 Introdução A Nutrição de PlantasDocumento24 páginasAula 1 Introdução A Nutrição de PlantasLarissa PortelaAinda não há avaliações
- Cobre - CuDocumento31 páginasCobre - CuI CAinda não há avaliações
- Nutrientes X Extrutura Da PlantaDocumento7 páginasNutrientes X Extrutura Da Plantaalinefarina.aAinda não há avaliações
- Trabalho Sobre CobreDocumento7 páginasTrabalho Sobre CobreLucas Pitangui do Prado Faria lpdpfAinda não há avaliações
- Capacidade do fungo Pleurotus ostreatus (cogumelo shimeji) na biorremediação de solos contaminados com chumbo (Pb)No EverandCapacidade do fungo Pleurotus ostreatus (cogumelo shimeji) na biorremediação de solos contaminados com chumbo (Pb)Ainda não há avaliações
- Glossário De Meio AmbienteNo EverandGlossário De Meio AmbienteAinda não há avaliações
- Biorremediação de Metais Pesados Utilizando Microalgas: Princípios e AplicaçõesNo EverandBiorremediação de Metais Pesados Utilizando Microalgas: Princípios e AplicaçõesAinda não há avaliações
- Aula 2 e 3 - Metabolismo Dos Carboidratos PDFDocumento48 páginasAula 2 e 3 - Metabolismo Dos Carboidratos PDFBibianaCristina Maestrello100% (1)
- Livro MicrobiologiaDocumento166 páginasLivro MicrobiologiaErica Viana100% (1)
- 4H-Ciclo de GlioxalatoDocumento9 páginas4H-Ciclo de Glioxalatoapi-27441481100% (5)
- Carboidratos: da ingestão à produção de energiaDocumento4 páginasCarboidratos: da ingestão à produção de energiaRonaldo ValmorbidaAinda não há avaliações
- Mitocôndrias: a organela celular responsável pela respiração e produção de energiaDocumento31 páginasMitocôndrias: a organela celular responsável pela respiração e produção de energiamamfm5Ainda não há avaliações
- 05 Perito Criminal EspecialDocumento12 páginas05 Perito Criminal EspecialIngride SantosAinda não há avaliações
- Gabarito Exercícios Metabolismo Energético PDFDocumento7 páginasGabarito Exercícios Metabolismo Energético PDFJoaquim Sixpence100% (1)
- NevrixDocumento2 páginasNevrixSOLDADO DIASSAinda não há avaliações
- FotossinteseDocumento8 páginasFotossinteseanon-309063100% (3)
- Importância Da Respiração CelularDocumento3 páginasImportância Da Respiração CelularAlexandra Duarte100% (1)
- Estudo Dirigido - Mitocôndrias e Cloroplastos.Documento3 páginasEstudo Dirigido - Mitocôndrias e Cloroplastos.Luiza LopesAinda não há avaliações
- Metabolismo de CarboidratosDocumento15 páginasMetabolismo de Carboidratosdani_eleine0% (1)
- PORFIRINAS e PORFÍRIAS: estrutura, biossíntese e regulaçãoDocumento55 páginasPORFIRINAS e PORFÍRIAS: estrutura, biossíntese e regulaçãoviinushkaAinda não há avaliações
- Apostila BiologiaDocumento41 páginasApostila BiologiaAndryo CardosoAinda não há avaliações
- O ciclo de Krebs e a produção de energiaDocumento6 páginasO ciclo de Krebs e a produção de energiaRichard WooliteAinda não há avaliações
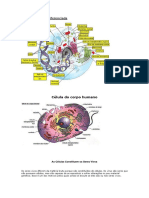
- Célula humana: estrutura e funçõesDocumento180 páginasCélula humana: estrutura e funçõesAna Paula Pereira Cunha100% (1)
- Resumo TopTopíssimo 2 PDFDocumento43 páginasResumo TopTopíssimo 2 PDFSofia SanchesAinda não há avaliações
- Vias Metabólicas No Jogo Da CapoeiraDocumento50 páginasVias Metabólicas No Jogo Da CapoeiraMessias Rodrigues100% (2)
- Sistema ATP-CP e glicólise anaeróbia no metabolismo muscularDocumento14 páginasSistema ATP-CP e glicólise anaeróbia no metabolismo muscularRicardo FurtadoAinda não há avaliações
- Produção de energia celular: glicólise, respiração e fermentaçãoDocumento10 páginasProdução de energia celular: glicólise, respiração e fermentaçãonomeAinda não há avaliações
- Resumo Oxidação de Ácidos GraxosDocumento7 páginasResumo Oxidação de Ácidos GraxosNicolle GabrielleAinda não há avaliações
- Ciclo de KrebsDocumento8 páginasCiclo de Krebsdielen23100% (1)
- 2º Teste de Bioenergética e Metabolismo Correcção 2Documento5 páginas2º Teste de Bioenergética e Metabolismo Correcção 2Carlos CostaAinda não há avaliações
- Aula Introd A MetabolismoDocumento95 páginasAula Introd A Metabolismoluci cristinaAinda não há avaliações
- Os principais tipos de açúcares e sua digestãoDocumento18 páginasOs principais tipos de açúcares e sua digestãoCamilla DiasAinda não há avaliações
- Respiração celular: glicólise e substratosDocumento21 páginasRespiração celular: glicólise e substratosDaniela ChavesAinda não há avaliações
- Bioenergetica PDFDocumento0 páginaBioenergetica PDFc4342893Ainda não há avaliações
- Bioenergética e MetabolismoDocumento47 páginasBioenergética e MetabolismoValeskaSenaAinda não há avaliações
- Oxidação dos ácidos graxos: via central de geração de energiaDocumento7 páginasOxidação dos ácidos graxos: via central de geração de energiaDariê CruvinelAinda não há avaliações
- Apostila de Biologia ENEM 2Documento7 páginasApostila de Biologia ENEM 2Ana030982Ainda não há avaliações