Você também pode gostar
- As 12 Camadas Da Personalidade Humana (Olavo de Carvalho)Documento17 páginasAs 12 Camadas Da Personalidade Humana (Olavo de Carvalho)Pisando Fofo100% (1)
- Como Parar de ProcrastinarDocumento8 páginasComo Parar de ProcrastinarVolki Lux100% (1)
- FIRE - Proibido e Perigoso - Jussara LealDocumento305 páginasFIRE - Proibido e Perigoso - Jussara LealRobson SodréAinda não há avaliações
- Unicamp Um Assunto Por QuestaoDocumento65 páginasUnicamp Um Assunto Por QuestaoLuiz WickAinda não há avaliações
- Linguagem - Cloze - Material QualitativoDocumento37 páginasLinguagem - Cloze - Material QualitativoTamires AlvesAinda não há avaliações
- Construção de Embarcações PequenasDocumento62 páginasConstrução de Embarcações PequenasHuascar Brito FeijoAinda não há avaliações
- Alimentos e Alimentação Animal: Maria Eduarda CabralDocumento75 páginasAlimentos e Alimentação Animal: Maria Eduarda CabraljhenniferAinda não há avaliações
- Guarulhos Manual Tratadores SPDocumento38 páginasGuarulhos Manual Tratadores SPAline AlcaláAinda não há avaliações
- 22técnicas de Projeção AstralDocumento27 páginas22técnicas de Projeção AstralAlisson PadilhaAinda não há avaliações
- AULA 01 - Introdução À PsicobiologiaDocumento29 páginasAULA 01 - Introdução À PsicobiologiadannhalabeAinda não há avaliações
- Apostila - Nutrição AnimalDocumento42 páginasApostila - Nutrição AnimalDenysson AmorimAinda não há avaliações
- 1 - U7 Ficha Formativa 1correcaoDocumento10 páginas1 - U7 Ficha Formativa 1correcaoMariana LopesAinda não há avaliações
- Aula 1 - Diferenças Anatômicas e Fisiológicas Entre Animais Ruminantes e Não-RuminantesDocumento24 páginasAula 1 - Diferenças Anatômicas e Fisiológicas Entre Animais Ruminantes e Não-RuminantesFrancislene Silveira SucupiraAinda não há avaliações
- Estrutura de Populações e Manejo FaunaDocumento6 páginasEstrutura de Populações e Manejo FaunaLaudineia MatiasAinda não há avaliações
- BubalinoculturaDocumento19 páginasBubalinoculturaLivia FerreiraAinda não há avaliações
- Icuria Dunensis ModelagemDocumento18 páginasIcuria Dunensis ModelagemAbilio Rafael Mucavele100% (1)
- Alimentação de Gatos Autor Archivaldo Reche Júnior e Marcela Malvini PimentaDocumento16 páginasAlimentação de Gatos Autor Archivaldo Reche Júnior e Marcela Malvini PimentaSimone StefaniAinda não há avaliações
- Aula 5. Bovinocultura - Nutrição de Ruminantes PDFDocumento76 páginasAula 5. Bovinocultura - Nutrição de Ruminantes PDFKely Vitória bezerra dos SantosAinda não há avaliações
- BioGeo10 - Teste 2Documento8 páginasBioGeo10 - Teste 2micaelamontezuma201150% (4)
- Alimentação e Nutrição de Organismos AquáticosDocumento57 páginasAlimentação e Nutrição de Organismos AquáticosJair Rastele100% (2)
- Aquário - Relatório FinalDocumento7 páginasAquário - Relatório FinalAna Caroline CasalvaraAinda não há avaliações
- Suinocultura Unidade II Noções de Nutrição e Alimentação de SuínosDocumento30 páginasSuinocultura Unidade II Noções de Nutrição e Alimentação de SuínosIsaura AliceAinda não há avaliações
- Atividade HemaglutinanteDocumento3 páginasAtividade HemaglutinanteJonathas DiegoAinda não há avaliações
- Avaliação de Métodos de Amostragem em Pastagens de Capim-Elefante (PennisetumDocumento8 páginasAvaliação de Métodos de Amostragem em Pastagens de Capim-Elefante (PennisetumHítalo RodriguesAinda não há avaliações
- Moreira Et Al. 2013 PDFDocumento9 páginasMoreira Et Al. 2013 PDFLV YoanyAinda não há avaliações
- Aminoacidos Felinos (Gatos)Documento11 páginasAminoacidos Felinos (Gatos)GUILHERME SOUTO DE SOUSAAinda não há avaliações
- Como Motivar Os Alunos e Professores Ns E.F.Documento675 páginasComo Motivar Os Alunos e Professores Ns E.F.profallex4438Ainda não há avaliações
- Template - Xviii - Sic - Final - Danilo 1Documento10 páginasTemplate - Xviii - Sic - Final - Danilo 1Danilo NunesAinda não há avaliações
- O Efeito de Probióticos Fúngicos Adicionados A Uma Dieta Rica em Grãos No Trato Gastrointestinal de OvinosDocumento13 páginasO Efeito de Probióticos Fúngicos Adicionados A Uma Dieta Rica em Grãos No Trato Gastrointestinal de OvinosKaio GabrielAinda não há avaliações
- Template - Xviii - Sic - Final - Danilo 1Documento10 páginasTemplate - Xviii - Sic - Final - Danilo 1Danilo NunesAinda não há avaliações
- Trabalho 1 HRPDocumento4 páginasTrabalho 1 HRPJamal Abdurabe AbdurabeAinda não há avaliações
- Digestão de Invertebrados 2Documento29 páginasDigestão de Invertebrados 2Roberto SantosAinda não há avaliações
- Endo e Ectoparasitas em JiboiasDocumento19 páginasEndo e Ectoparasitas em Jiboiasabraaobiologo2Ainda não há avaliações
- Métodos Titulométricos (Preciptação e Neutralização)Documento25 páginasMétodos Titulométricos (Preciptação e Neutralização)LEANDRO BORGESAinda não há avaliações
- Manejo, Reabilitacao e Soltura de Mamiferos SelvagensDocumento6 páginasManejo, Reabilitacao e Soltura de Mamiferos Selvagenslilianelionco4407Ainda não há avaliações
- Kami Export - Richard Silva - QUESTÕES BIOLOGIA AULA 4Documento3 páginasKami Export - Richard Silva - QUESTÕES BIOLOGIA AULA 4Richard SantosAinda não há avaliações
- Degradabilidade Ruminal e Digestibilidade Intestinal de Alimentos PorDocumento9 páginasDegradabilidade Ruminal e Digestibilidade Intestinal de Alimentos PorDelizio Marcanizzy DBAinda não há avaliações
- FINAL - DANILO - Correção 1Documento10 páginasFINAL - DANILO - Correção 1Danilo NunesAinda não há avaliações
- Projeto PibicDocumento12 páginasProjeto PibicAndreza SantosAinda não há avaliações
- Alevinos Carpa Capim Forragem e RaçãoDocumento5 páginasAlevinos Carpa Capim Forragem e RaçãoTatudoherado TthAinda não há avaliações
- Desempenho de Coelhos Alimentados Com Farinha de Tenebrio MolitorDocumento8 páginasDesempenho de Coelhos Alimentados Com Farinha de Tenebrio MolitorANDRESSA SANTANNA NATELAinda não há avaliações
- Seminários PT 2 - Sara GrecovDocumento5 páginasSeminários PT 2 - Sara GrecovSara GrecovAinda não há avaliações
- Artigo Suínos e Felinos - Nutrição de MonogástricosDocumento5 páginasArtigo Suínos e Felinos - Nutrição de MonogástricosJessica da Conceicao SantosAinda não há avaliações
- Coleta de FezesDocumento9 páginasColeta de FezesFabrício Eugênio AraújoAinda não há avaliações
- Coeficiente de DigestibilidadeDocumento8 páginasCoeficiente de DigestibilidadeDaniel ArrudaAinda não há avaliações
- Nitrogenado Rumin MonoDocumento10 páginasNitrogenado Rumin MonoLuiz AnastacioAinda não há avaliações
- FISIOLOGIA DA DIGESTÃO E ABSORÇÃO DE CARBOIDRATOS NAS AVES Renato - Furlan PDFDocumento18 páginasFISIOLOGIA DA DIGESTÃO E ABSORÇÃO DE CARBOIDRATOS NAS AVES Renato - Furlan PDFJuliana AlexandreAinda não há avaliações
- Biofisiologiacomparada07102011livro 140722002101 Phpapp02Documento122 páginasBiofisiologiacomparada07102011livro 140722002101 Phpapp02Mateus MatosAinda não há avaliações
- 1055 3929 1 PBDocumento6 páginas1055 3929 1 PBHelder RezendeAinda não há avaliações
- Cactos TraduzidoDocumento7 páginasCactos Traduzidojose.pauloAinda não há avaliações
- Avaliacao Parasitologica de Ratos (Rattus Novergicus)Documento7 páginasAvaliacao Parasitologica de Ratos (Rattus Novergicus)wagneramadoAinda não há avaliações
- Apresentação NutriçãoDocumento5 páginasApresentação NutriçãoJosefa PereiraAinda não há avaliações
- Let 5803Documento3 páginasLet 5803antoniohomsiAinda não há avaliações
- 110-122 - Rev - FCT (1) - 9Documento13 páginas110-122 - Rev - FCT (1) - 9Hélio Årlindo ManhiqueAinda não há avaliações
- Daphnia OrvalhinhaDocumento2 páginasDaphnia OrvalhinhaFilipe BarrosoAinda não há avaliações
- CONSTRUÇÃO E CARACTERIZAÇÃO DE UMA BIBLIOTECA GENÔMICA ENRIQUECIDA EM MICROSSATÉLITES PARA O APAPÁ (Pellona Castelnaeana, PRISTIGASTERIDAE)Documento3 páginasCONSTRUÇÃO E CARACTERIZAÇÃO DE UMA BIBLIOTECA GENÔMICA ENRIQUECIDA EM MICROSSATÉLITES PARA O APAPÁ (Pellona Castelnaeana, PRISTIGASTERIDAE)Sica UfpaAinda não há avaliações
- Valor Nutritivo Das Pastagens TropicaisDocumento15 páginasValor Nutritivo Das Pastagens TropicaisLeonardo SouzaAinda não há avaliações
- Aula 1 - Origem Da Zootecnia PDFDocumento17 páginasAula 1 - Origem Da Zootecnia PDFCharles MagnumAinda não há avaliações
- Trabalho Fisiologia Ii EspecialDocumento29 páginasTrabalho Fisiologia Ii EspecialMaria Danieli PiresAinda não há avaliações
- Cópia de Efeito Das Lectinas Nos Insetos SchistocercaDocumento5 páginasCópia de Efeito Das Lectinas Nos Insetos SchistocercaGabriela FernandesAinda não há avaliações
- Apostila AlimentosDocumento49 páginasApostila AlimentosfosquimaAinda não há avaliações
- 662-Texto Do Artigo-3523-1-10-20200804Documento8 páginas662-Texto Do Artigo-3523-1-10-20200804Carlos SaboiaAinda não há avaliações
- Alga Marrom (Ascophyllum Nodosum) para Alevinos de Tilápia Do NiloDocumento8 páginasAlga Marrom (Ascophyllum Nodosum) para Alevinos de Tilápia Do NiloJosué ItaloAinda não há avaliações
- Revista10, v.10, N.2aa.p65Documento9 páginasRevista10, v.10, N.2aa.p65Carlos TheoAinda não há avaliações
- Comportamento Ingestivo de Caprinos Moxotó e Canindé Submetidos À Alimentação À Vontade e RestritaDocumento8 páginasComportamento Ingestivo de Caprinos Moxotó e Canindé Submetidos À Alimentação À Vontade e Restritafrancisconeto95Ainda não há avaliações
- Slides ResumoDocumento6 páginasSlides ResumoEverton SantosAinda não há avaliações
- Botanica Aquatica DigitalDocumento6 páginasBotanica Aquatica DigitalEcodrop AquapaisagismoAinda não há avaliações
- Trabalho Sobre Cadeia Alimetar ISCEDDocumento16 páginasTrabalho Sobre Cadeia Alimetar ISCEDmussa nassordino nassordinoAinda não há avaliações
- Avaliação Do Comportamento de Cutias Dasyprocta Azarae e Dasyprocta Leporina (Rodentia Dasyproctidae) em Cativeiro e SemicativeiroDocumento15 páginasAvaliação Do Comportamento de Cutias Dasyprocta Azarae e Dasyprocta Leporina (Rodentia Dasyproctidae) em Cativeiro e SemicativeiroJani PereiraAinda não há avaliações
- Retinol na Produção In Vitro de Embriões CaprinosNo EverandRetinol na Produção In Vitro de Embriões CaprinosAinda não há avaliações
- III Asrochas Arquivosquerelatamahistriadaterra 101001102128 Phpapp02 PDFDocumento38 páginasIII Asrochas Arquivosquerelatamahistriadaterra 101001102128 Phpapp02 PDF300 BaltaAinda não há avaliações
- 10 Ano Geologia A Terra e Os Seus Subsistemas em Interaco Parte 1 1222283956046808 9 PDFDocumento20 páginas10 Ano Geologia A Terra e Os Seus Subsistemas em Interaco Parte 1 1222283956046808 9 PDF300 BaltaAinda não há avaliações
- 1-Extincao Dinossauros PDFDocumento7 páginas1-Extincao Dinossauros PDF300 BaltaAinda não há avaliações
- Re Sumo RochasDocumento20 páginasRe Sumo RochasAna Carla Pereira BarbosaAinda não há avaliações
- Captura de Ecrã 2021-04-27 À(s) 19.10.25 PDFDocumento14 páginasCaptura de Ecrã 2021-04-27 À(s) 19.10.25 PDF300 BaltaAinda não há avaliações
- Rochas Sedimentares 1223035143979212 9 PDFDocumento31 páginasRochas Sedimentares 1223035143979212 9 PDF300 BaltaAinda não há avaliações
- Visita de Estudo Relatã Rio Baltazar 3Documento11 páginasVisita de Estudo Relatã Rio Baltazar 3300 BaltaAinda não há avaliações
- Captura de Ecrã 2021-04-16 À(s) 10.56.42 PDFDocumento6 páginasCaptura de Ecrã 2021-04-16 À(s) 10.56.42 PDF300 BaltaAinda não há avaliações
- Primeiro PeriodoDocumento4 páginasPrimeiro Periodo300 BaltaAinda não há avaliações
- Aprender PortuguêsDocumento12 páginasAprender Português300 BaltaAinda não há avaliações
- Etica Moral e Direito No Transumanismo eDocumento20 páginasEtica Moral e Direito No Transumanismo eDeicon Davi Siqueira PiresAinda não há avaliações
- Trapoeraba, Erva de Difícil Controle em CafezaisDocumento2 páginasTrapoeraba, Erva de Difícil Controle em CafezaisEmmanuel CecchettoAinda não há avaliações
- Trabalho de BiologiaDocumento20 páginasTrabalho de BiologiaManuela AquinoAinda não há avaliações
- Guia Completo GBC LIFE Mar21Documento135 páginasGuia Completo GBC LIFE Mar21Elton ElvesAinda não há avaliações
- Formigas (Apresentação)Documento15 páginasFormigas (Apresentação)Rubens MMaiaAinda não há avaliações
- Sistema Digestório de RuminantesDocumento18 páginasSistema Digestório de RuminantesLorrayne BorgesAinda não há avaliações
- Semana 07Documento190 páginasSemana 07BlochMeisterAinda não há avaliações
- Aciocinio Logico para Concursos - 4a EdDocumento12 páginasAciocinio Logico para Concursos - 4a EdMariana Salles Franco TorresAinda não há avaliações
- Lista Nucleo Celular, Ciclo Celular e MitoseDocumento3 páginasLista Nucleo Celular, Ciclo Celular e MitoseAna Paula RodriguesAinda não há avaliações
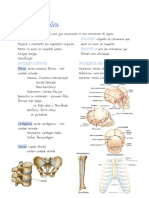
- ArticulaçõesDocumento3 páginasArticulaçõesEllen Gonçalves MoreiraAinda não há avaliações
- Currículo LattesDocumento5 páginasCurrículo LattesThiagoAinda não há avaliações
- Prova 3 Ano - GenéticaDocumento2 páginasProva 3 Ano - GenéticaAyane DantasAinda não há avaliações
- A Invencao Da InfanciaDocumento11 páginasA Invencao Da InfanciaPaticene Fone PaticeneAinda não há avaliações
- Princípios de Neurociências KANDEL - 5 EDDocumento16 páginasPrincípios de Neurociências KANDEL - 5 EDMARCOS DIASAinda não há avaliações
- Letra XXDocumento17 páginasLetra XXSandra VicenteAinda não há avaliações
- Simulação Clonagem MolecularDocumento84 páginasSimulação Clonagem MolecularGRAZIELLE DUARTE SILVAAinda não há avaliações
- Universidade Do Estado Do Rio de Janeiro: Hotel Van EetveldeDocumento24 páginasUniversidade Do Estado Do Rio de Janeiro: Hotel Van EetveldeMariana ValenteAinda não há avaliações
- Vantagens Definitivo Versão CompletissimaDocumento29 páginasVantagens Definitivo Versão CompletissimafrantiescoAinda não há avaliações
- Resumo Bioquímica Do Tecido Nervoso - Kayla 154Documento4 páginasResumo Bioquímica Do Tecido Nervoso - Kayla 154kayladalva81Ainda não há avaliações