Você também pode gostar
- Sistema Cardiovascular 9ºDocumento24 páginasSistema Cardiovascular 9ºExplica-meArouca0% (1)
- Rejuvenescer Com O Plasma Sanguíneo Dos JovensNo EverandRejuvenescer Com O Plasma Sanguíneo Dos JovensAinda não há avaliações
- E-Book L-PRF e Stick BoneDocumento22 páginasE-Book L-PRF e Stick BoneMac Prado Alves Mac100% (2)
- Aula 5-12-04 - 2022 Biofísica - UnipDocumento47 páginasAula 5-12-04 - 2022 Biofísica - Uniplauana RebecaAinda não há avaliações
- Ciclo Celular: Divisão e Reprodução das CélulasDocumento13 páginasCiclo Celular: Divisão e Reprodução das CélulasArthur AngeloAinda não há avaliações
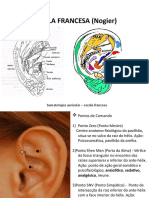
- Escola Francesa de AuriculoterapiaDocumento15 páginasEscola Francesa de AuriculoterapiaRoberta Higina KoralekAinda não há avaliações
- Apostila Do Curso Tratamento de Feridas e CurativosDocumento74 páginasApostila Do Curso Tratamento de Feridas e CurativosBeatriz Santos100% (1)
- Ciclo CelularDocumento49 páginasCiclo CelularIsaura Chiyeck100% (1)
- MitoseDocumento8 páginasMitosekadunetoAinda não há avaliações
- Manual de Formação CelularDocumento16 páginasManual de Formação CelularMafalda RamosAinda não há avaliações
- Aula 4 - Ciclo Celular Mitose e Meiose - Aspectos GenéticosDocumento30 páginasAula 4 - Ciclo Celular Mitose e Meiose - Aspectos GenéticosAdryelle Mendes100% (1)
- O ciclo celular e seu controleDocumento5 páginasO ciclo celular e seu controlemoematimoAinda não há avaliações
- 3 - O Ciclo de Divisão CelularDocumento13 páginas3 - O Ciclo de Divisão CelularEduardo MatosAinda não há avaliações
- Ciclo Celular e PontosDocumento6 páginasCiclo Celular e PontosRafaela CostaAinda não há avaliações
- Ciclo celular: interfase, mitose e regulaçãoDocumento22 páginasCiclo celular: interfase, mitose e regulaçãoAna CanastraAinda não há avaliações
- Genetica IiDocumento11 páginasGenetica IiAmaralAinda não há avaliações
- IntroduçãoDocumento26 páginasIntroduçãojotapinto22Ainda não há avaliações
- M1P2Documento13 páginasM1P2Hemily RodriguesAinda não há avaliações
- Ciclo CelularDocumento7 páginasCiclo CelularMatheus ToledoAinda não há avaliações
- Ciclo CelularDocumento10 páginasCiclo CelularMiguel Alberto LinoAinda não há avaliações
- Ciclo Celular e o Sistema de ControleDocumento4 páginasCiclo Celular e o Sistema de ControleGabriela LeãoAinda não há avaliações
- Ciclo Celular - JalekoDocumento21 páginasCiclo Celular - JalekoÉrica LageAinda não há avaliações
- Ciclo Celularbiologia2Documento16 páginasCiclo Celularbiologia2ana santosAinda não há avaliações
- O ciclo celular: compreendendo as fases da mitose e a importância da regulação do processoDocumento8 páginasO ciclo celular: compreendendo as fases da mitose e a importância da regulação do processovelascofonsecaAinda não há avaliações
- Ciclo celular: multiplicação, diferenciação e regulação do processoDocumento11 páginasCiclo celular: multiplicação, diferenciação e regulação do processoLuccaAinda não há avaliações
- Resumo P4Documento20 páginasResumo P4LuccaAinda não há avaliações
- O MISTERIOSO CORPO HUMANODocumento11 páginasO MISTERIOSO CORPO HUMANOEdmario De Jesus CorreiaAinda não há avaliações
- Octavia-Textos NormativosDocumento10 páginasOctavia-Textos NormativosArmando TamboAinda não há avaliações
- Ciclo CelularDocumento4 páginasCiclo CelularVero GuañunaAinda não há avaliações
- Ciclo Celular e ApoptoseDocumento6 páginasCiclo Celular e ApoptoseAna CristinaAinda não há avaliações
- (Alberts, 6 Ed) Cap. 17 - Ciclo CelularDocumento14 páginas(Alberts, 6 Ed) Cap. 17 - Ciclo CelularDenis NascimentoAinda não há avaliações
- Ciclo Celular e MitoseDocumento18 páginasCiclo Celular e MitoseBruna SilvaAinda não há avaliações
- Compreendendo os pontos de checagem do ciclo celular e o reparo do DNADocumento12 páginasCompreendendo os pontos de checagem do ciclo celular e o reparo do DNAWalterAinda não há avaliações
- Citologia, Histologia e GenéticaDocumento14 páginasCitologia, Histologia e Genéticafatimalima.nutriAinda não há avaliações
- Ciclo Celular e Suas FasesDocumento10 páginasCiclo Celular e Suas FasesLarissa Batista Carvalho SantosAinda não há avaliações
- Biologia 2Documento16 páginasBiologia 2Aluno Alice da Silva CalhauAinda não há avaliações
- Ciclo celular - Fases e regulaçãoDocumento8 páginasCiclo celular - Fases e regulaçãojoanaAinda não há avaliações
- Student 53572Documento6 páginasStudent 53572LuisNemGamerAinda não há avaliações
- Ciclo Celular: Interfase e MitoseDocumento12 páginasCiclo Celular: Interfase e MitoseLuizAinda não há avaliações
- A Divisão Celular e o Ciclo CelularDocumento6 páginasA Divisão Celular e o Ciclo CelularLorena CostaAinda não há avaliações
- Ciclo Celular - Cerca 2º AnoDocumento9 páginasCiclo Celular - Cerca 2º AnoJosé Pedro Moreirão Cerca Monteiro de OliveiraAinda não há avaliações
- Divisão Celular-Mitose e Meiose 1Documento55 páginasDivisão Celular-Mitose e Meiose 1Marineide AmorimAinda não há avaliações
- Ciclo celular, fases da mitose e sistema de controleDocumento5 páginasCiclo celular, fases da mitose e sistema de controleForwomenAinda não há avaliações
- Bio Celular II (1) - 022317 - 120206Documento99 páginasBio Celular II (1) - 022317 - 120206domindoma28Ainda não há avaliações
- Ciclo CelularDocumento11 páginasCiclo Celularmanueljpseabra3587Ainda não há avaliações
- Biologia 3Documento44 páginasBiologia 3sem limitesAinda não há avaliações
- Pontos de Checagem Do Ciclo CelularDocumento6 páginasPontos de Checagem Do Ciclo CelularAline GranatoAinda não há avaliações
- Trabalho de Pesquisa Ciclo CelularDocumento8 páginasTrabalho de Pesquisa Ciclo CelularHonoria KiwaAinda não há avaliações
- Reguladores do ciclo celular: ciclinas, Cdks e APC/CDocumento6 páginasReguladores do ciclo celular: ciclinas, Cdks e APC/CTúlio HenriqueAinda não há avaliações
- Ciclo Vegetal_125421Documento10 páginasCiclo Vegetal_125421joxdenzel977Ainda não há avaliações
- O ciclo celular: as etapas da divisão celularDocumento7 páginasO ciclo celular: as etapas da divisão celularInes GomesAinda não há avaliações
- Aula Ciclo, Mitose e Meiose PDFDocumento52 páginasAula Ciclo, Mitose e Meiose PDFfabilleneAinda não há avaliações
- Ciclo Celular: o Que É, Etapas, Controle, Resumo - Biologia NetDocumento3 páginasCiclo Celular: o Que É, Etapas, Controle, Resumo - Biologia NetlamdcoelhoAinda não há avaliações
- Tics Isis Ciclo CelularDocumento2 páginasTics Isis Ciclo CelularMaria DanieleAinda não há avaliações
- Regulação do ciclo celular eucariotaDocumento41 páginasRegulação do ciclo celular eucariotaJoão Frederico MusialAinda não há avaliações
- Estudo de Caso 2 - Guilherme Jesus Da SilvaDocumento8 páginasEstudo de Caso 2 - Guilherme Jesus Da SilvaeuguisilvajAinda não há avaliações
- Ciclo CelularDocumento9 páginasCiclo CelularViviane Carvalho SuethAinda não há avaliações
- Ciclo CelularDocumento6 páginasCiclo CelularMariana DiasAinda não há avaliações
- BiologiaDocumento8 páginasBiologiaRita RodriguesAinda não há avaliações
- Ciclo Celular e Mitose 2021 Com VídeosDocumento22 páginasCiclo Celular e Mitose 2021 Com VídeosBruna Nogueira de AraujoAinda não há avaliações
- Documento 1Documento8 páginasDocumento 1Taremy Gody69Ainda não há avaliações
- Documento Sem NomeDocumento17 páginasDocumento Sem NomeMatilde MendesAinda não há avaliações
- Resumo Ciclo CelularDocumento8 páginasResumo Ciclo CelularBruna KelletAinda não há avaliações
- Divisão celular e ciclo celularDocumento22 páginasDivisão celular e ciclo celularFabio Gonçalves Da SilvaAinda não há avaliações
- Citoesqueleto - Maria Gabriela Franco de LimaDocumento11 páginasCitoesqueleto - Maria Gabriela Franco de LimaMaria Gabriela franco de limaAinda não há avaliações
- ApresentaçãoDocumento32 páginasApresentaçãoMaria Gabriela franco de limaAinda não há avaliações
- Corantes e Aditivos Alimentares (Final)Documento38 páginasCorantes e Aditivos Alimentares (Final)Maria Gabriela franco de limaAinda não há avaliações
- SeminárioDocumento33 páginasSeminárioMaria Gabriela franco de limaAinda não há avaliações
- Yoga benefícios corpo menteDocumento2 páginasYoga benefícios corpo menteAna MariaAinda não há avaliações
- Aula 4 - Distúrbios Estéticos (Alopécia Androgênica) PDFDocumento44 páginasAula 4 - Distúrbios Estéticos (Alopécia Androgênica) PDFChristhyanne Dos Santos CarreiroAinda não há avaliações
- Tabela TubosDocumento1 páginaTabela TubosCad VetAinda não há avaliações
- O potencial de membrana em neurôniosDocumento26 páginasO potencial de membrana em neurôniosJéssica LeãoAinda não há avaliações
- Exercício de Neuroanatomia PRONTODocumento4 páginasExercício de Neuroanatomia PRONTOFernando MaiaAinda não há avaliações
- Lista de Teses e Dissertações do ICBDocumento39 páginasLista de Teses e Dissertações do ICBFilipe Camera de BonaAinda não há avaliações
- Sistema Sensorial - StryerDocumento31 páginasSistema Sensorial - StryerLucas Ferreira Gama MarquiniAinda não há avaliações
- Apostila Neurologia (Prova 1)Documento36 páginasApostila Neurologia (Prova 1)João Marcos Pereira Neiva RodriguesAinda não há avaliações
- Treinamento sensório-motorDocumento101 páginasTreinamento sensório-motorDoomerAinda não há avaliações
- Tecido CartilaginosoDocumento27 páginasTecido CartilaginosoMuryllo PossamaiAinda não há avaliações
- Cólica EquinaDocumento7 páginasCólica Equinaalejr896Ainda não há avaliações
- PP28 Célula - Unidade Estrutural e Funcional (Alunos)Documento60 páginasPP28 Célula - Unidade Estrutural e Funcional (Alunos)Gustavo LapoAinda não há avaliações
- Morfologia LeucocitáriaDocumento105 páginasMorfologia Leucocitáriaray_limaaAinda não há avaliações
- ECG Como Você Nunca Viu - Capítulo 1Documento23 páginasECG Como Você Nunca Viu - Capítulo 1Izamara SouzaAinda não há avaliações
- Biologia - Tipos de TecidosDocumento3 páginasBiologia - Tipos de TecidosJoao VictorAinda não há avaliações
- Mapa Mental - EnzimasDocumento1 páginaMapa Mental - EnzimasAna BeaAinda não há avaliações
- Exercícios - 2 Lei de Mendel e Grupos SanguíneosDocumento2 páginasExercícios - 2 Lei de Mendel e Grupos SanguíneosDaniel JatobaAinda não há avaliações
- Sistema Tegumentar: Estrutura e Funções da Pele e AnexosDocumento25 páginasSistema Tegumentar: Estrutura e Funções da Pele e AnexosLarissa Sena100% (1)
- Definição de hipóxia e hipoxemiaDocumento2 páginasDefinição de hipóxia e hipoxemiaJhuanna Carolina EcheveriaAinda não há avaliações
- Fisioterapia Dermato-Funcional: Mais que beleza, saúdeDocumento6 páginasFisioterapia Dermato-Funcional: Mais que beleza, saúdeJanneAinda não há avaliações
- Fisiologia do SNC e SNP emDocumento50 páginasFisiologia do SNC e SNP emVanderlan FranciscoAinda não há avaliações
- Estudo DirigidoDocumento9 páginasEstudo DirigidoStephany Summaya AmorimAinda não há avaliações
- 001.resumo - Sistema Circulatório - Coração e VasosDocumento10 páginas001.resumo - Sistema Circulatório - Coração e VasosNanami LinAinda não há avaliações
- Roteiro OSTEOLOGIADocumento7 páginasRoteiro OSTEOLOGIANayche TortatoAinda não há avaliações
- Estudo Dirigido I Anatomia Vegetal I Semestre Letivo de 2016 ProfDocumento8 páginasEstudo Dirigido I Anatomia Vegetal I Semestre Letivo de 2016 Prof202310469Ainda não há avaliações