Você também pode gostar
- Sistema nervosoDocumento5 páginasSistema nervosokalianetnAinda não há avaliações
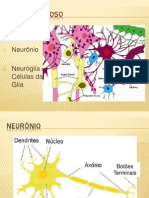
- Tecido NervosoDocumento8 páginasTecido NervosoAnne KellyAinda não há avaliações
- Guia de Referência Do Sistema Nervoso - DIGITALDocumento136 páginasGuia de Referência Do Sistema Nervoso - DIGITALnaemi ishiguroAinda não há avaliações
- Tecido NervosoDocumento83 páginasTecido NervosoSérgio SilasAinda não há avaliações
- Sistema NervosoDocumento81 páginasSistema NervosoGuilherme RebliAinda não há avaliações
- Tecido nervoso: estrutura, funções e tipos de célulasDocumento16 páginasTecido nervoso: estrutura, funções e tipos de célulasYwlla ReisAinda não há avaliações
- SISTEMA NERVOSO 2024Documento58 páginasSISTEMA NERVOSO 2024José Augusto Casagrande CorreiaAinda não há avaliações
- Sistema nervoso: estrutura e funçõesDocumento9 páginasSistema nervoso: estrutura e funçõesLeo RamosAinda não há avaliações
- Tecido NervosoDocumento6 páginasTecido Nervosomariana machadoAinda não há avaliações
- O Sistema Nervoso e seus ComponentesDocumento20 páginasO Sistema Nervoso e seus ComponentesDiego da Rosa100% (1)
- Neuroanatomia e Sistema NervosoDocumento9 páginasNeuroanatomia e Sistema NervosoCaio OliveiraAinda não há avaliações
- SNC e tipos de neurôniosDocumento19 páginasSNC e tipos de neurôniosPaibra Diana100% (1)
- O tecido nervoso e seus componentesDocumento87 páginasO tecido nervoso e seus componentesChay FerreiraAinda não há avaliações
- Fisioterapia NeurologiaDocumento63 páginasFisioterapia NeurologiaAne SilvaAinda não há avaliações
- ESTUDO DIRIGIDO - AV1 - NeuroanatomofisiologiaDocumento7 páginasESTUDO DIRIGIDO - AV1 - Neuroanatomofisiologiatamara.guimab2Ainda não há avaliações
- Histologia Do Sistema NervosoDocumento60 páginasHistologia Do Sistema NervosoMax Nunes100% (1)
- Organização celular do sistema nervosoDocumento3 páginasOrganização celular do sistema nervosoGlória MamédioAinda não há avaliações
- Tecido NervosoDocumento3 páginasTecido NervosoMarcos Antonio D'Queiroz JuniorAinda não há avaliações
- Generalidades Do Tecido NervosoDocumento4 páginasGeneralidades Do Tecido Nervosofabiofranzoni123Ainda não há avaliações
- 2 - Tecido NervosoDocumento6 páginas2 - Tecido NervosoLuciano PeixotoAinda não há avaliações
- Sistema NervosoDocumento4 páginasSistema NervosoKanasa 03Ainda não há avaliações
- Sistema Nervoso AnatomiaDocumento24 páginasSistema Nervoso AnatomiaRafaela RochaAinda não há avaliações
- Resumo Tecido NervosoDocumento5 páginasResumo Tecido NervosoRaiane ValentimAinda não há avaliações
- Tecido Nervoso 2Documento3 páginasTecido Nervoso 2Domingos EuroAinda não há avaliações
- Células Nervosas e Divisão do Sistema NervosoDocumento8 páginasCélulas Nervosas e Divisão do Sistema NervosoWesley Ramalho SouzaAinda não há avaliações
- Tecido nervoso AULA 4Documento29 páginasTecido nervoso AULA 4fgamaisaAinda não há avaliações
- Sistema Nervoso-1-36Documento36 páginasSistema Nervoso-1-36Kaio VictorAinda não há avaliações
- NervosoDocumento11 páginasNervosoLucas Brumatti SetubalAinda não há avaliações
- Anatomia Resumo DigitadoDocumento8 páginasAnatomia Resumo DigitadoSaúde Da MulherAinda não há avaliações
- NeurofisiologiaDocumento12 páginasNeurofisiologiaMaria HelenaAinda não há avaliações
- Tutoria 2 - Sistema NervosoDocumento14 páginasTutoria 2 - Sistema Nervoso002-025384Ainda não há avaliações
- Aula Neurofisiologia Primeiro Bimestre 2024.1Documento56 páginasAula Neurofisiologia Primeiro Bimestre 2024.1leitaolucileydeAinda não há avaliações
- Anotações Livro NeuroanatomiaDocumento6 páginasAnotações Livro NeuroanatomiaCaroline SlongoAinda não há avaliações
- NEUROFISIOLOGIADocumento10 páginasNEUROFISIOLOGIAjofrepereira29Ainda não há avaliações
- Sistema NervosoDocumento4 páginasSistema NervosoAntonio BernardoAinda não há avaliações
- Neurônios e células da glia fundamentais para o SNDocumento7 páginasNeurônios e células da glia fundamentais para o SNSimone AndreozziAinda não há avaliações
- Neuroanatomia EaD 2022Documento23 páginasNeuroanatomia EaD 2022Laura GarciaAinda não há avaliações
- Sistema Nervoso: tecido nervoso e potencial de açãoDocumento48 páginasSistema Nervoso: tecido nervoso e potencial de açãoSueli LemosAinda não há avaliações
- Aula 1 - Introdução Ao Sistema Nervoso NovaDocumento32 páginasAula 1 - Introdução Ao Sistema Nervoso NovaAnne GlauzerAinda não há avaliações
- Fisiologia Do Sistema NervosoDocumento41 páginasFisiologia Do Sistema NervosothalysonamorinAinda não há avaliações
- parte escrita do trabalho de histologiaDocumento4 páginasparte escrita do trabalho de histologiasuyaneAinda não há avaliações
- Tecido Nervoso: Neurônios, Neuróglia e MielinaDocumento34 páginasTecido Nervoso: Neurônios, Neuróglia e MielinadevidelimaAinda não há avaliações
- Sistema NervosoDocumento3 páginasSistema NervosoJérisson BarrosAinda não há avaliações
- Introdução Ao Sistema NervosoDocumento6 páginasIntrodução Ao Sistema NervosoAnne GabrielleAinda não há avaliações
- NeurociênciasDocumento6 páginasNeurociênciasAmanda YaniAinda não há avaliações
- Tecido NervosoDocumento48 páginasTecido NervosoCristiane Faustina de CarvalhoAinda não há avaliações
- 4.sistema NervosoDocumento38 páginas4.sistema NervosoTiago MachadoAinda não há avaliações
- Sistema Nervoso e Principais DoençasDocumento38 páginasSistema Nervoso e Principais DoençasBEATRIZ BARBOSAAinda não há avaliações
- Aula 2 - Introdução Ao Estudo Da NafDocumento43 páginasAula 2 - Introdução Ao Estudo Da NafMarcílio CésarAinda não há avaliações
- Anatomia-Sistema NervosoDocumento93 páginasAnatomia-Sistema NervosojackeAinda não há avaliações
- Apostila Fisiologia Do Sistema NervosoDocumento73 páginasApostila Fisiologia Do Sistema NervosoRogerio BarancelliAinda não há avaliações
- NOVA APOSTILA - Sistema NervosoDocumento102 páginasNOVA APOSTILA - Sistema NervosoOtávio LageAinda não há avaliações
- Sistema NervosoDocumento15 páginasSistema NervosoYasmin SpínolaAinda não há avaliações
- Bases morfológicas e funcionais do sistema nervosoDocumento44 páginasBases morfológicas e funcionais do sistema nervosoFelipe BrunoAinda não há avaliações
- Sistema Nervoso VertebradosDocumento4 páginasSistema Nervoso VertebradosernestoAinda não há avaliações
- Resumo de Generalidades Do Sistema NervosoDocumento16 páginasResumo de Generalidades Do Sistema NervosomelissadsmAinda não há avaliações
- Abordagem Morfofuncional Do Sistema NervosoDocumento69 páginasAbordagem Morfofuncional Do Sistema NervosoBeatriz RochaAinda não há avaliações
- Sistema nervoso: estrutura anatômica e tecido nervosoDocumento15 páginasSistema nervoso: estrutura anatômica e tecido nervosokéssiaAinda não há avaliações
- Histologia Tecido NervosoDocumento12 páginasHistologia Tecido NervosomarianyAinda não há avaliações
- Dicionário de BrandingDocumento2 páginasDicionário de BrandingIanaraTeixeiraAinda não há avaliações
- Computação I - Python Laboratório 1 funçõesDocumento2 páginasComputação I - Python Laboratório 1 funçõesFabricioAinda não há avaliações
- Detox Diet GuideDocumento26 páginasDetox Diet GuideMarcelo MirandaAinda não há avaliações
- Análise Do Poema - Ode Triunfal - de Álvaro de CamposDocumento5 páginasAnálise Do Poema - Ode Triunfal - de Álvaro de CamposFrancisco MesquitaAinda não há avaliações
- Lazarillo de TormesDocumento30 páginasLazarillo de TormesBeatriz VasconcelosAinda não há avaliações
- Processo N.º 23-18-L (Erro de Direito)Documento3 páginasProcesso N.º 23-18-L (Erro de Direito)Miguel Augusto PmfAinda não há avaliações
- O Narcisismo No Contexto Da Maternidade: Algumas Evidências EmpíricasDocumento8 páginasO Narcisismo No Contexto Da Maternidade: Algumas Evidências EmpíricasPaulo ConnorAinda não há avaliações
- Jogo Dos CanudosDocumento2 páginasJogo Dos CanudosAlexandre NegraoAinda não há avaliações
- PHP 5, MySQL 5 e Ajax para desenvolvimento webDocumento166 páginasPHP 5, MySQL 5 e Ajax para desenvolvimento webFabio MattesAinda não há avaliações
- Arte e TecnologiaDocumento6 páginasArte e TecnologiaMagrãoAinda não há avaliações
- Unidade 4Documento6 páginasUnidade 4Raphael FerreiraAinda não há avaliações
- Lista 6 ABC e Custeio VariávelDocumento5 páginasLista 6 ABC e Custeio Variávelroneisantos2011Ainda não há avaliações
- Amor e CuidadoDocumento28 páginasAmor e CuidadomarcosagnerAinda não há avaliações
- Reinos de Taifas e o rei poeta Almu'tamidDocumento2 páginasReinos de Taifas e o rei poeta Almu'tamidNetfixe LolAinda não há avaliações
- Caderno Dos EsportesDocumento1 páginaCaderno Dos EsportesRonald RegisAinda não há avaliações
- Atividade FuncaoDocumento9 páginasAtividade FuncaoHelena MiekoAinda não há avaliações
- Aula 31 - 03 Out - MasterDocumento27 páginasAula 31 - 03 Out - MasterJoão VictorAinda não há avaliações
- Livro PAISAGEMDocumento258 páginasLivro PAISAGEMCamilla Rodrigues100% (1)
- Aula 1 - Introdução As Doenças Crônicas - A Realidade Do BrasilDocumento10 páginasAula 1 - Introdução As Doenças Crônicas - A Realidade Do BrasilVitor Silva LimaAinda não há avaliações
- 5ef PDFDocumento29 páginas5ef PDFCleunice AlmeidaAinda não há avaliações
- Paes e Paula - Repensando Os Estudos OrganizacDocumento298 páginasPaes e Paula - Repensando Os Estudos OrganizacElisabete Stradiotto SiqueiraAinda não há avaliações
- Dodf 026 06-02-2023 Integra-80-82Documento3 páginasDodf 026 06-02-2023 Integra-80-82Marc ArnoldiAinda não há avaliações
- Plano de ensino de Psicodiagnóstico (2022.2Documento7 páginasPlano de ensino de Psicodiagnóstico (2022.2João Lucas TeixeiraAinda não há avaliações
- Apac EditavelDocumento1 páginaApac Editavelcirurgia hrs88% (17)
- Kant e o idealismo na Crítica da Razão PuraDocumento20 páginasKant e o idealismo na Crítica da Razão PuraIsmael De Oliveira GerolamoAinda não há avaliações
- Projeto Alimentação SaúdavelDocumento3 páginasProjeto Alimentação SaúdavelPriscila NascimentoAinda não há avaliações
- Protocolo de EndodontiaDocumento17 páginasProtocolo de EndodontiaMauricio DiasAinda não há avaliações
- Edital SelecaoDocumento16 páginasEdital SelecaoHenrique Eduardo Guarani-kaiowáAinda não há avaliações
- Jogos Divertidos Com Imagens PDFDocumento71 páginasJogos Divertidos Com Imagens PDFElton Robert Vieira PachecoAinda não há avaliações
- Saúde PúblicaDocumento43 páginasSaúde Públicajoaca Galvão100% (3)
- Série Didática para o Apoio a Formação de Professores de Química: Volume 2: MoléculasNo EverandSérie Didática para o Apoio a Formação de Professores de Química: Volume 2: MoléculasNota: 5 de 5 estrelas5/5 (1)
- Equilíbrio hormonal _ Recupere equilíbrio hormonal, libido, sono e emagreça já!No EverandEquilíbrio hormonal _ Recupere equilíbrio hormonal, libido, sono e emagreça já!Nota: 5 de 5 estrelas5/5 (2)
- Uma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNo EverandUma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNota: 4 de 5 estrelas4/5 (13)
- Atlas de anatomia: Imagens humanas e veterináriasNo EverandAtlas de anatomia: Imagens humanas e veterináriasNota: 4.5 de 5 estrelas4.5/5 (2)
- Equilíbrio iônico: aplicações em química analíticaNo EverandEquilíbrio iônico: aplicações em química analíticaAinda não há avaliações
- Práticas de Genética, Biologia Molecular, Biotecnologia e EvoluçãoNo EverandPráticas de Genética, Biologia Molecular, Biotecnologia e EvoluçãoNota: 4 de 5 estrelas4/5 (1)
- Simetria Molecular III: Série Didática para o apoio a formação de professores de QuímicaNo EverandSimetria Molecular III: Série Didática para o apoio a formação de professores de QuímicaAinda não há avaliações
- Problemas de Cinética e Reatores Químicos: 100 Problemas Resolvidos, 500 Problemas Propostos (Com Respostas)No EverandProblemas de Cinética e Reatores Químicos: 100 Problemas Resolvidos, 500 Problemas Propostos (Com Respostas)Ainda não há avaliações
- Química da Cerveja: Uma Abordagem Química e Bioquímica das Matérias-Primas, Processo de Produção e da Composição dos Compostos de Sabores da CervejaNo EverandQuímica da Cerveja: Uma Abordagem Química e Bioquímica das Matérias-Primas, Processo de Produção e da Composição dos Compostos de Sabores da CervejaAinda não há avaliações
- Uma breve história da química: Da alquimia às ciências moleculares modernasNo EverandUma breve história da química: Da alquimia às ciências moleculares modernasAinda não há avaliações
- Para Gostar de Química: Introdução à Química GeralNo EverandPara Gostar de Química: Introdução à Química GeralAinda não há avaliações
- Físico-química de Blendas e Compósitos PoliméricosNo EverandFísico-química de Blendas e Compósitos PoliméricosNota: 1 de 5 estrelas1/5 (1)
- Detetive Por Um Dia: Aprendendo A Investigar Por Meio Da Química ForenseNo EverandDetetive Por Um Dia: Aprendendo A Investigar Por Meio Da Química ForenseAinda não há avaliações
- Uso do Policloreto de Alumínio (PAC): vantagens sobre o Sulfato de Alumínio em sistema de tratamento de águaNo EverandUso do Policloreto de Alumínio (PAC): vantagens sobre o Sulfato de Alumínio em sistema de tratamento de águaAinda não há avaliações
- Vigilância Sanitária Nos Estados E MunicípiosNo EverandVigilância Sanitária Nos Estados E MunicípiosAinda não há avaliações