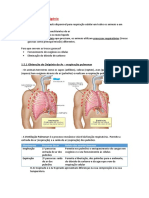
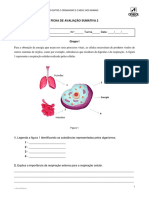
Você também pode gostar
- TeoricoDocumento20 páginasTeoricoLaraAinda não há avaliações
- 1 BioquimicaDocumento18 páginas1 BioquimicaCleane Carvalho Dos SantosAinda não há avaliações
- FISIOLOGIA Termorregulação, Osmorregulação e ExcreçãoDocumento28 páginasFISIOLOGIA Termorregulação, Osmorregulação e Excreçãoingridrakuel123Ainda não há avaliações
- TeoricoDocumento28 páginasTeoricoEmmanuel VasconcelosAinda não há avaliações
- MAPA - Material de Avaliação Prática Da Aprendizagem: DisciplinaDocumento6 páginasMAPA - Material de Avaliação Prática Da Aprendizagem: Disciplinajose carlos felizAinda não há avaliações
- Respiracao CelularDocumento16 páginasRespiracao CelularDercioAinda não há avaliações
- Poluição Atmosferica PDFDocumento102 páginasPoluição Atmosferica PDFAnaClaudiaEnfAinda não há avaliações
- Fundamentos Da Fisiologia ComparadaDocumento26 páginasFundamentos Da Fisiologia Comparadaingridrakuel123Ainda não há avaliações
- Medicina Abdominal em Cães e GatosDocumento129 páginasMedicina Abdominal em Cães e GatosClaudia SilvaAinda não há avaliações
- Composições Do Ar - 071402Documento2 páginasComposições Do Ar - 071402Rosildo Brasil PintoAinda não há avaliações
- Teorico 2Documento20 páginasTeorico 2joao victorAinda não há avaliações
- Bio Celular e MolecularDocumento40 páginasBio Celular e MolecularNataly MarquesAinda não há avaliações
- FAC - Questões 1Documento3 páginasFAC - Questões 1Juliana TavaresAinda não há avaliações
- RESPIRACAODocumento40 páginasRESPIRACAOfiorirs2007Ainda não há avaliações
- Fisiologia - Respiração Interna e ExternaDocumento7 páginasFisiologia - Respiração Interna e Externacb_penatrujillo100% (4)
- Resumi Concluido PDFDocumento19 páginasResumi Concluido PDFFrancisco Rosario JaimeAinda não há avaliações
- Biopráticas: Atividades experimentaisNo EverandBiopráticas: Atividades experimentaisAinda não há avaliações
- Cadeia Alimentar e Ciclos Biogeoquímicos 1ºDocumento9 páginasCadeia Alimentar e Ciclos Biogeoquímicos 1ºMário Luiz DalmolinAinda não há avaliações
- Bioquimica Estrutural e Metabolica 4Documento32 páginasBioquimica Estrutural e Metabolica 4Estética Mel FerrettiAinda não há avaliações
- Mapa - Biologia e BioquimicaDocumento4 páginasMapa - Biologia e BioquimicaRonaldo ValmorbidaAinda não há avaliações
- UntitledDocumento8 páginasUntitledEdman MoldeAinda não há avaliações
- Bioquímica - GeralDocumento156 páginasBioquímica - Geralmadeseb740Ainda não há avaliações
- Ciclos Biogeoquimicos - ResumoDocumento4 páginasCiclos Biogeoquimicos - ResumobrunoAinda não há avaliações
- Prova de Biologia 3° Bimestre - 2A e 2B - GABDocumento2 páginasProva de Biologia 3° Bimestre - 2A e 2B - GABCarla LimaAinda não há avaliações
- Relatório Compartimentalização CelularDocumento4 páginasRelatório Compartimentalização CelularRianna Ornella Almeida SobralAinda não há avaliações
- FisiologiaDocumento10 páginasFisiologiaIamara Policarpo100% (2)
- RelatorioDocumento9 páginasRelatorioAlayde SimedoAinda não há avaliações
- Aula 01Documento15 páginasAula 01Jhilliane RibeiroAinda não há avaliações
- 2010 Volume 1 CADERNODOALUNO QUIMICA EnsinoMedio 3aserie Caderno Do AlunoDocumento48 páginas2010 Volume 1 CADERNODOALUNO QUIMICA EnsinoMedio 3aserie Caderno Do AlunoRones DiasAinda não há avaliações
- Química OrgânicaDocumento24 páginasQuímica OrgânicaLaianeantunesAinda não há avaliações
- Lab Bio Cel TeciDocumento26 páginasLab Bio Cel TeciGaius Cassius LonginusAinda não há avaliações
- AULA - Metabolismo EnergéticoDocumento9 páginasAULA - Metabolismo EnergéticoIsadora GusmãoAinda não há avaliações
- Ciclos BiogeoquímicosDocumento23 páginasCiclos BiogeoquímicospaulobuenoAinda não há avaliações
- BiologiaDocumento96 páginasBiologiaraf4el4Ainda não há avaliações
- Mapa India BiologiaDocumento5 páginasMapa India Biologiajeder santanaAinda não há avaliações
- Unidader 1Documento22 páginasUnidader 1Tony TanikagiAinda não há avaliações
- Atividade Bio 08-04Documento4 páginasAtividade Bio 08-04Luiz Gustavo De Macedo FreitasAinda não há avaliações
- Fundamentos Da BioquimicaDocumento120 páginasFundamentos Da BioquimicaCarol ValençaAinda não há avaliações
- TeoricoDocumento28 páginasTeoricoLaraAinda não há avaliações
- CadernoDoAluno 2014 Vol1 Baixa CN Quimica EM 3SDocumento98 páginasCadernoDoAluno 2014 Vol1 Baixa CN Quimica EM 3SDouglas Ricardo de AssisAinda não há avaliações
- Unidade IV - CarboidratosDocumento18 páginasUnidade IV - CarboidratosStanislav Ozzy PetrovAinda não há avaliações
- Teorico IVDocumento28 páginasTeorico IVLeonardo LimaAinda não há avaliações
- Camila SanchesDocumento12 páginasCamila SanchesMarieAinda não há avaliações
- Biofisica Aplicada A Biomedicina 1Documento42 páginasBiofisica Aplicada A Biomedicina 1Estética Mel FerrettiAinda não há avaliações
- Relatório de Biologia - Taxa Metabólica de Peixes e LâminasDocumento9 páginasRelatório de Biologia - Taxa Metabólica de Peixes e LâminasRodolfo FerreiraAinda não há avaliações
- Química Orgânica 1Documento24 páginasQuímica Orgânica 1LaianeantunesAinda não há avaliações
- Relatório QG Unip - EduardoDocumento15 páginasRelatório QG Unip - Eduardoleonidas GamerAinda não há avaliações
- Tecnologia Dos Alimentos TeoricoDocumento24 páginasTecnologia Dos Alimentos TeoricoMilly MoreiraAinda não há avaliações
- BIOQUIMICADocumento78 páginasBIOQUIMICALorena Henriques100% (2)
- RESUMODocumento8 páginasRESUMOEstudos VitóriaAinda não há avaliações
- Roteiro de PesquisaDocumento2 páginasRoteiro de PesquisaLucileia garciaAinda não há avaliações
- Ebook Desco Top Semana 01 Turma JulhoDocumento421 páginasEbook Desco Top Semana 01 Turma JulhoHenrique PadãoAinda não há avaliações
- Ciclos BiogeoquimicosDocumento12 páginasCiclos BiogeoquimicosPaula AlmeidaAinda não há avaliações
- Bases Macromoleculares Da Constituição CelularDocumento8 páginasBases Macromoleculares Da Constituição CelularIsac Francisco RaulAinda não há avaliações
- V GowinDocumento4 páginasV GowinX100% (1)
- Impressão - Texto Substâncias Inorgânicas e Orgênicas Dos Seres VivosDocumento4 páginasImpressão - Texto Substâncias Inorgânicas e Orgênicas Dos Seres VivosKelma Cristina de FreitasAinda não há avaliações
- Biologia - Namimatsu - Apostila1Documento10 páginasBiologia - Namimatsu - Apostila1Ciencias PPT100% (1)
- Ginástica respiratória (Traduzido): Purificação - Saúde - Força - EnergiaNo EverandGinástica respiratória (Traduzido): Purificação - Saúde - Força - EnergiaAinda não há avaliações
- Comportamento Animal: Uma Introdução aos Métodos e à Ecologia ComportamentalNo EverandComportamento Animal: Uma Introdução aos Métodos e à Ecologia ComportamentalNota: 3 de 5 estrelas3/5 (1)
- Ficha-Informativa-Animais-Vertebrados-e-Invertebrados 3º AnoDocumento3 páginasFicha-Informativa-Animais-Vertebrados-e-Invertebrados 3º Anocarpraia0% (1)
- Ciencias Naturais Módulo 1 1.5 Obtenção de Oxigénio Do Ar e Da AguaDocumento4 páginasCiencias Naturais Módulo 1 1.5 Obtenção de Oxigénio Do Ar e Da AguaAna PintoAinda não há avaliações
- 5CN6 - Respiração CelularDocumento3 páginas5CN6 - Respiração CelularmariaconceiaoAinda não há avaliações
- Morfologia e Composição Do Pescado - 1729Documento55 páginasMorfologia e Composição Do Pescado - 1729Marta AzevedoAinda não há avaliações
- Fisiologia Animal ComparadaDocumento135 páginasFisiologia Animal ComparadaCaroline Silva Alves100% (2)
- Resumo de Ciencias Naturais 6 Ano - Fonte O Bichinho Do SaberDocumento157 páginasResumo de Ciencias Naturais 6 Ano - Fonte O Bichinho Do SaberMónica JanuárioAinda não há avaliações
- Ficha Informativa - OsmorregulaçãoDocumento8 páginasFicha Informativa - OsmorregulaçãoMaria Clara GregoAinda não há avaliações
- Zoologia - Peixes - (109 Questões)Documento69 páginasZoologia - Peixes - (109 Questões)Aldemir GabrielAinda não há avaliações
- 06.PPt OrgaosRespAnimaisDocumento30 páginas06.PPt OrgaosRespAnimaisBeatriz SantosAinda não há avaliações
- Ficha Informativa Animais Vertebrados e InvertebradosDocumento3 páginasFicha Informativa Animais Vertebrados e Invertebradossjfwn8n788% (8)
- Peixes - Trabalho de Biologia (Doc. Oficial)Documento9 páginasPeixes - Trabalho de Biologia (Doc. Oficial)Nikolas RochaAinda não há avaliações
- Especialidade de TubarõesDocumento11 páginasEspecialidade de TubarõesMariana SampaioAinda não há avaliações
- Excretor ReprodutorDocumento40 páginasExcretor ReprodutorRosaVinhalAinda não há avaliações
- Biologia Dos Vertebrados PDFDocumento64 páginasBiologia Dos Vertebrados PDFFCiências100% (4)
- Resumo Ciências 6º AnoDocumento12 páginasResumo Ciências 6º AnoLuis DiasAinda não há avaliações
- ApostilaDocumento27 páginasApostilaEduardo ViolaAinda não há avaliações
- Resumo 10 - Unidade III - Transformação e Utilização de Energia Pelos Seres VivosDocumento8 páginasResumo 10 - Unidade III - Transformação e Utilização de Energia Pelos Seres VivosGlennda Jorge PinheiroAinda não há avaliações
- Resumo Zoologia Wbio 2 PDFDocumento3 páginasResumo Zoologia Wbio 2 PDFChrystiane AlmeidaAinda não há avaliações
- EC - 7º Ano - Ciências - Animais VertebradosDocumento11 páginasEC - 7º Ano - Ciências - Animais VertebradosRosana ValeAinda não há avaliações
- Aparelho Respiratório Dos VertebradosDocumento31 páginasAparelho Respiratório Dos Vertebradosrafrancoo75% (4)
- V GowinDocumento4 páginasV GowinX100% (1)
- Trocas Gasosas - Biologia e Geologia 10º AnoDocumento34 páginasTrocas Gasosas - Biologia e Geologia 10º AnoCarlota FiguerolaAinda não há avaliações
- 3 Anfibios - Dom Bosco - CursoDocumento5 páginas3 Anfibios - Dom Bosco - CursoMarcos Antonio SantosAinda não há avaliações
- Lista Exercicios.Documento2 páginasLista Exercicios.Francieli de JesusAinda não há avaliações
- Ficha de Avaliação - Sist. RespiratórioDocumento6 páginasFicha de Avaliação - Sist. RespiratórioSandra CardosoAinda não há avaliações
- AD2 - Zoologia - Tayane AbsDocumento4 páginasAD2 - Zoologia - Tayane AbsRaíssa ManzouqueAinda não há avaliações
- 2 Bimestre Aula 2. Classe Crustacea PDFDocumento24 páginas2 Bimestre Aula 2. Classe Crustacea PDFFlorinda Marques SilvaAinda não há avaliações
- Lista de Exercícios InvertebradosDocumento9 páginasLista de Exercícios InvertebradosValquiria SilvaAinda não há avaliações
- Resumos de Biologia e GeologiaDocumento34 páginasResumos de Biologia e GeologiaMariana MeninoAinda não há avaliações
- Ae ccn6 Teste 2Documento7 páginasAe ccn6 Teste 2XandraAinda não há avaliações