Você também pode gostar
- Potencial Elétrico ResumoDocumento2 páginasPotencial Elétrico ResumoIasmim MaiaraAinda não há avaliações
- Diferenças Entre Benzodiazepínicos e BarbitúricosDocumento2 páginasDiferenças Entre Benzodiazepínicos e BarbitúricosAntonella Marreiros100% (1)
- Paratormônio, Calcitriol e CalcitoninaDocumento2 páginasParatormônio, Calcitriol e CalcitoninaFernanda Hermeto100% (1)
- Apostila Urinário PDFDocumento14 páginasApostila Urinário PDFEllison Silva AraújoAinda não há avaliações
- Capitulo 6 Resumo Fisiologia GuytonDocumento5 páginasCapitulo 6 Resumo Fisiologia Guytonmuller maiaAinda não há avaliações
- Litiase UrinariaDocumento5 páginasLitiase UrinariaRadiologia VelosoAinda não há avaliações
- Aula 4 - Potencial de Membrana e AçãoDocumento14 páginasAula 4 - Potencial de Membrana e AçãoRafael LopesAinda não há avaliações
- Potencial de AçãoDocumento28 páginasPotencial de AçãoArthur Calandrine100% (1)
- 31 Metabolismo ÓsseoDocumento10 páginas31 Metabolismo ÓsseoFábio Marcelino Duarte DuarteAinda não há avaliações
- RADIOLOGIA 01 - Introdução - MED RESUMOS (JAN-2012)Documento0 páginaRADIOLOGIA 01 - Introdução - MED RESUMOS (JAN-2012)Priscila FerroAinda não há avaliações
- Resumo Cap 38 Guyton - Bruno HollandaDocumento2 páginasResumo Cap 38 Guyton - Bruno HollandaGustavo TagliaviniAinda não há avaliações
- EF Aparelhourogenital APSDocumento3 páginasEF Aparelhourogenital APSEN Ferreira40% (5)
- Sistema Renina Angiotensina AldosteronaDocumento8 páginasSistema Renina Angiotensina AldosteronaÍbis Ariana100% (1)
- Sindrome Do Tunel Do CarpoDocumento19 páginasSindrome Do Tunel Do Carpoapi-3857420100% (2)
- Embriologia CardiacaDocumento25 páginasEmbriologia Cardiacaapi-3700467100% (5)
- Questões - Anatomia Da Pelve FemininaDocumento3 páginasQuestões - Anatomia Da Pelve FemininaandréAinda não há avaliações
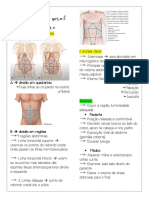
- Exame Fisico AbdominalDocumento7 páginasExame Fisico AbdominalAna Clara GardioliAinda não há avaliações
- Reposição HidroeletrolíticaDocumento13 páginasReposição HidroeletrolíticaJonathas Dias Melo100% (1)
- Fisiologia Renal - GuytonDocumento4 páginasFisiologia Renal - GuytonCristine Gondim Maia100% (4)
- Fraturas Cervicais Altas 2018Documento110 páginasFraturas Cervicais Altas 2018marilia leaoAinda não há avaliações
- Resumo Cap 27 Guyton Bruno HollandaDocumento5 páginasResumo Cap 27 Guyton Bruno HollandaJoão Vitor Assmann Fontana100% (1)
- Sistema Nervoso SimpáticoDocumento11 páginasSistema Nervoso SimpáticoMateus Dos Santos BandeiraAinda não há avaliações
- MicrocirculaçãoDocumento31 páginasMicrocirculaçãoReinaldo RiquetAinda não há avaliações
- Fibrose CísticaDocumento21 páginasFibrose Císticacarol ferrazAinda não há avaliações
- Atividade Contextualizada de Anatomia HumanaDocumento2 páginasAtividade Contextualizada de Anatomia HumanaAdilson De Oliveira Silva100% (1)
- Resposta A Broncodilatador em EspirometriaDocumento20 páginasResposta A Broncodilatador em EspirometriaPaulo Gurgel Carlos da Silva100% (1)
- 10 - Fisiologia RenalDocumento20 páginas10 - Fisiologia RenalSauloMachadoAinda não há avaliações
- Cetoacidose Diabética HijpiiDocumento1 páginaCetoacidose Diabética Hijpiiwolve100% (1)
- Exercício - TelencéfaloDocumento14 páginasExercício - TelencéfaloLuciana LoboAinda não há avaliações
- Estudo Dirigido-Sistema Genital Masculino.Documento5 páginasEstudo Dirigido-Sistema Genital Masculino.Cristina MeloAinda não há avaliações
- Resumo de EcgDocumento21 páginasResumo de EcgdgmoellerAinda não há avaliações
- Fibrilação Atrial PDFDocumento3 páginasFibrilação Atrial PDFPedro EliasAinda não há avaliações
- Função RenalDocumento31 páginasFunção Renalherbertpsf100% (3)
- Cap 19 GuytonDocumento2 páginasCap 19 GuytonmaxbufaAinda não há avaliações
- Resumo de Farmacologia Sobre AntihipertensivosDocumento6 páginasResumo de Farmacologia Sobre AntihipertensivosNatan Mello100% (2)
- Câncer de PênisDocumento3 páginasCâncer de PênisSylviahMasanto100% (1)
- HAS - Hipertensão Arterial SistêmicaDocumento18 páginasHAS - Hipertensão Arterial SistêmicaAline Grana100% (1)
- Farmacologia Do Sistema VascularDocumento14 páginasFarmacologia Do Sistema VascularLorena Moreira VieiraAinda não há avaliações
- Slides de Crise HipertensivaDocumento31 páginasSlides de Crise HipertensivaJéssica Tavares100% (1)
- Sintese e Liberação Da AcetilcolinaDocumento1 páginaSintese e Liberação Da AcetilcolinaSuelen IzabelAinda não há avaliações
- Resumo de BioeletrogeneseDocumento15 páginasResumo de BioeletrogeneseBruno Avelar100% (1)
- Capitulo 32 GuytonDocumento7 páginasCapitulo 32 GuytonRenan Helmer Noelves0% (1)
- ResumoMembranaCelular 1557739734476 1571484171Documento6 páginasResumoMembranaCelular 1557739734476 1571484171Carlos MonturilAinda não há avaliações
- 1 Pancreatite AgudaDocumento89 páginas1 Pancreatite AgudaDean JamesAinda não há avaliações
- Med Resumos - Fisiologia Da RespiraçãoDocumento12 páginasMed Resumos - Fisiologia Da RespiraçãoEsther Beiral100% (3)
- 3 - Resumo Eduardo - Semiologia Do TóraxDocumento21 páginas3 - Resumo Eduardo - Semiologia Do TóraxJulianna Lutz100% (1)
- Exame Torácico e Dos PulmõesDocumento11 páginasExame Torácico e Dos PulmõesSilvaleideAtaidesAinda não há avaliações
- Bioquímica Clínica - Wagner PintoDocumento935 páginasBioquímica Clínica - Wagner Pintodaniele frotaAinda não há avaliações
- Questões-Sistema-Nervoso-Autonômico (Resolvida)Documento3 páginasQuestões-Sistema-Nervoso-Autonômico (Resolvida)Leon FernandesAinda não há avaliações
- Cardiopatia HipertensivaDocumento10 páginasCardiopatia HipertensivaSarah Lima CamposAinda não há avaliações
- Inflamação AgudaDocumento4 páginasInflamação Agudaapi-3797560100% (7)
- Apostila Teorica RenalDocumento23 páginasApostila Teorica RenalHenrique MacetãoAinda não há avaliações
- Semiologia Do Aparelho Osteomuscular - 2014Documento22 páginasSemiologia Do Aparelho Osteomuscular - 2014Luiz Geraldo100% (1)
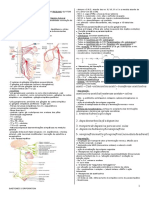
- Medresumos 2014 Anatomia Humana Sistmica 10 Sistema GenitalDocumento14 páginasMedresumos 2014 Anatomia Humana Sistmica 10 Sistema GenitalDenise PereiraAinda não há avaliações
- Capitulo 8 Reumo Fisiologia GuytonDocumento4 páginasCapitulo 8 Reumo Fisiologia Guytonmuller maia100% (1)
- Fechamento Problema 3Documento7 páginasFechamento Problema 3anna luisaAinda não há avaliações
- Sinalização Elétrica Dos NeurôniosDocumento6 páginasSinalização Elétrica Dos NeurôniosJúliaAinda não há avaliações
- Sistema Nervoso (Aula 1)Documento14 páginasSistema Nervoso (Aula 1)talitabenvindoAinda não há avaliações
- Potencial de AçãoDocumento6 páginasPotencial de AçãoRafael ReisAinda não há avaliações
- Tema 2 - Comunicação Neural, Sentidos Somatossensoriais e Sentidos EspeciaisDocumento25 páginasTema 2 - Comunicação Neural, Sentidos Somatossensoriais e Sentidos EspeciaisFlávia Couto LealAinda não há avaliações
- Afecções Neuroftalmológicas ModificadoDocumento12 páginasAfecções Neuroftalmológicas ModificadoRoberto Almeida DolceAinda não há avaliações
- Prevenção Da Cegueira Infantil ModificadoDocumento10 páginasPrevenção Da Cegueira Infantil Modificadomuller maiaAinda não há avaliações
- Estrabismo ModificadoDocumento10 páginasEstrabismo Modificadomuller maiaAinda não há avaliações
- Ametropias ModificadoDocumento11 páginasAmetropias Modificadomuller maiaAinda não há avaliações
- Ametropias ModificadoDocumento11 páginasAmetropias Modificadomuller maiaAinda não há avaliações
- Teste Do Reflexo VermelhoDocumento6 páginasTeste Do Reflexo Vermelhomuller maiaAinda não há avaliações
- Prevenção Da Cegueira Infantil ModificadoDocumento10 páginasPrevenção Da Cegueira Infantil Modificadomuller maiaAinda não há avaliações
- Relatório 4 - Contagem Diferencial de LeucócitosDocumento1 páginaRelatório 4 - Contagem Diferencial de Leucócitosmuller maiaAinda não há avaliações
- Apostila ImunologiaDocumento21 páginasApostila Imunologiamuller maiaAinda não há avaliações
- Relatório 1 - Esfregaço SanguíneoDocumento1 páginaRelatório 1 - Esfregaço Sanguíneomuller maiaAinda não há avaliações
- Relatório 2 - Células SanguíneasDocumento1 páginaRelatório 2 - Células Sanguíneasmuller maiaAinda não há avaliações
- Relatório 3 - Órgãos LinfoidesDocumento2 páginasRelatório 3 - Órgãos Linfoidesmuller maiaAinda não há avaliações
- Potencial de Membrana e Potenciais de AçaoDocumento6 páginasPotencial de Membrana e Potenciais de Açaomuller maiaAinda não há avaliações
- Sifilis - Manual Aula 2 PDFDocumento23 páginasSifilis - Manual Aula 2 PDFEloi MenezesAinda não há avaliações
- Capítulo 55 - Controle Cortical e Do Tronco Cerebral Da Função Motora - 4 PáginasDocumento4 páginasCapítulo 55 - Controle Cortical e Do Tronco Cerebral Da Função Motora - 4 PáginasandreamokadaAinda não há avaliações
- Capítulo 45 - Organização Do SNC, Funções Básicas Das Sinapses e Neurotransmissores - 6 PágDocumento6 páginasCapítulo 45 - Organização Do SNC, Funções Básicas Das Sinapses e Neurotransmissores - 6 PágandreamokadaAinda não há avaliações
- Resumo Guyton (Cap. 60)Documento3 páginasResumo Guyton (Cap. 60)muller maiaAinda não há avaliações
- Captulo 56 Guyton Contribuies Do Cerebelo e Dos Ncleos Da Base para o Controle Motor GlobalDocumento3 páginasCaptulo 56 Guyton Contribuies Do Cerebelo e Dos Ncleos Da Base para o Controle Motor Globalmuller maiaAinda não há avaliações
- Resumo Guyton Vol. 11 (Caps. 01 A 05)Documento20 páginasResumo Guyton Vol. 11 (Caps. 01 A 05)Jones Correa100% (1)
- Capítulo 48 - Sensações Somáticas Ii - Dor, Cefaléia e Sensações Térmicas - 3 PáginasDocumento3 páginasCapítulo 48 - Sensações Somáticas Ii - Dor, Cefaléia e Sensações Térmicas - 3 Páginasandreamokada100% (1)
- Capitulo 46 Resumo Fisiologia GuytonDocumento6 páginasCapitulo 46 Resumo Fisiologia Guytonmuller maia100% (1)
- Capitulo 55 Controles Cortical e Do Tronco Cerebral Na Funçao MotoraDocumento2 páginasCapitulo 55 Controles Cortical e Do Tronco Cerebral Na Funçao Motoramuller maia100% (1)
- Capitulo 48 Resumo Fisiologia GuytonDocumento8 páginasCapitulo 48 Resumo Fisiologia Guytonmuller maiaAinda não há avaliações
- Capitulo 8 Reumo Fisiologia GuytonDocumento4 páginasCapitulo 8 Reumo Fisiologia Guytonmuller maia100% (1)
- Capitulo 45 Resumo Fisiologia GuytonDocumento9 páginasCapitulo 45 Resumo Fisiologia Guytonmuller maiaAinda não há avaliações
- Capitulo 7 Resumo Fisiologia GuytonDocumento4 páginasCapitulo 7 Resumo Fisiologia Guytonmuller maiaAinda não há avaliações
- Capitulo 8 Reumo Fisiologia GuytonDocumento4 páginasCapitulo 8 Reumo Fisiologia Guytonmuller maia100% (1)
- Capitulo 9 Resumo Fisiologia GuytonDocumento7 páginasCapitulo 9 Resumo Fisiologia Guytonmuller maiaAinda não há avaliações
- Bulário Sinóptico PDFDocumento34 páginasBulário Sinóptico PDFtokashiAinda não há avaliações
- 1.SBV (Aha) - 2020Documento29 páginas1.SBV (Aha) - 2020AntonioAinda não há avaliações
- Planejamento de Aula-TECIDOS - Docx 1Documento5 páginasPlanejamento de Aula-TECIDOS - Docx 1Catarina GuelerAinda não há avaliações
- Pele e Anexos Ajuste25!10!11Documento45 páginasPele e Anexos Ajuste25!10!11Polly Carlos MachadoAinda não há avaliações
- Disciplina de Ortodontia Preventiva: Movimento Dentário Ortodôntico - Parte 2Documento28 páginasDisciplina de Ortodontia Preventiva: Movimento Dentário Ortodôntico - Parte 2Daroana FerrareseAinda não há avaliações
- Sistema Respiratório - WEB ACADDocumento56 páginasSistema Respiratório - WEB ACADFranciel StuelpAinda não há avaliações
- ChlorophytaDocumento2 páginasChlorophytaRicardo JoppertAinda não há avaliações
- Apostila - DPOCDocumento5 páginasApostila - DPOCDiego CoelhoAinda não há avaliações
- Hormônios VegetaisDocumento6 páginasHormônios VegetaisMarianaAinda não há avaliações
- Ventosa AulaDocumento50 páginasVentosa AulaFisiobinho100% (6)
- Morfoanatomia de Folhas e Caules de Passiflora EdulisDocumento17 páginasMorfoanatomia de Folhas e Caules de Passiflora EdulisNaira SoaresAinda não há avaliações
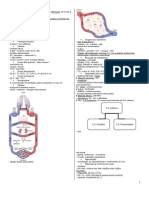
- Sistema CardiovascularDocumento24 páginasSistema CardiovascularThayná CrisóstomoAinda não há avaliações
- Sistema Nervoso e EndocrinoDocumento15 páginasSistema Nervoso e EndocrinoJoão Pedro CarvalhoAinda não há avaliações
- (Histologia) Sistema UrinárioDocumento18 páginas(Histologia) Sistema UrinárioSofia VilarAinda não há avaliações
- Resolução 4º Teste CN 6º AnoDocumento7 páginasResolução 4º Teste CN 6º AnoJoão Maria DiasAinda não há avaliações
- Aula 1 - Introdução A Formas de Liberação de FármacosDocumento189 páginasAula 1 - Introdução A Formas de Liberação de FármacosSuzana Bender0% (1)
- Membrana Plasmatica SlidesDocumento36 páginasMembrana Plasmatica SlidesSheila Cassenotte Ferreira100% (1)
- Mapa Mental AmiodaronaDocumento1 páginaMapa Mental AmiodaronaEdna100% (1)
- Estruturas e Funções - HIPOTÁLAMO, SUBTÁLAMO, EPITÁLAMO, TÁLAMODocumento10 páginasEstruturas e Funções - HIPOTÁLAMO, SUBTÁLAMO, EPITÁLAMO, TÁLAMORosana Carvalho de Souza100% (1)
- O Quarteto Fantástico Da FelicidadeDocumento2 páginasO Quarteto Fantástico Da FelicidadeArmando Ribeiro100% (1)
- Apostila Aula 1Documento14 páginasApostila Aula 1Vagner EliseuAinda não há avaliações
- Prática - Avaliação Da ATM PDFDocumento5 páginasPrática - Avaliação Da ATM PDFJoílton OliveiraAinda não há avaliações
- Anatomia Humana Enfermagem Sistema CardiovascularDocumento207 páginasAnatomia Humana Enfermagem Sistema CardiovascularVitor Sousa de nascimentoAinda não há avaliações
- Prova de EspecialidadesDocumento9 páginasProva de EspecialidadesBruno FellipeAinda não há avaliações
- AxtaxantinaDocumento2 páginasAxtaxantinaCarlos AcguariusAinda não há avaliações
- Exercicios 2Documento6 páginasExercicios 2Pedro Neves25% (4)
- Microagulhamento Como Agente PotencializadorDocumento5 páginasMicroagulhamento Como Agente PotencializadorandrelalAinda não há avaliações
- Asfixia PerinatalDocumento8 páginasAsfixia PerinatalmarquesconceicaoAinda não há avaliações
- Herança Dos Grupos Sanguíneos Do Sistema ABODocumento11 páginasHerança Dos Grupos Sanguíneos Do Sistema ABOJuliana SousaAinda não há avaliações
- ATIVIDADE AV1 FisioepsicoDocumento4 páginasATIVIDADE AV1 FisioepsicoRaphael AmorimAinda não há avaliações