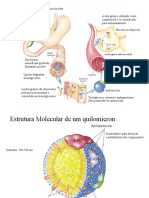
Você também pode gostar
- Aula 9 - Integração MetabólicaDocumento6 páginasAula 9 - Integração MetabólicaArielle MoraesAinda não há avaliações
- Estudo DirigidoDocumento16 páginasEstudo DirigidoRenatoMatias100% (2)
- 3 Resumo Fotossintese 2020 21Documento9 páginas3 Resumo Fotossintese 2020 21Laura MonteiroAinda não há avaliações
- Metabolismo Energético PDFDocumento10 páginasMetabolismo Energético PDFEu Sou BobinhoAinda não há avaliações
- Estudo Dirigido 1 - Bioquímica - Janeiro 2023Documento6 páginasEstudo Dirigido 1 - Bioquímica - Janeiro 2023AMANDA GLÓRIA100% (1)
- Resumo Bioquímica CarboidratosDocumento7 páginasResumo Bioquímica CarboidratosIana MagalhãesAinda não há avaliações
- Via Das Pentoses FosfatoDocumento1 páginaVia Das Pentoses FosfatoMarion SoaresAinda não há avaliações
- Questões de BioquimicaDocumento7 páginasQuestões de BioquimicaPatrícia AlmeidaAinda não há avaliações
- Aminoácidos e Proteínas PDFDocumento5 páginasAminoácidos e Proteínas PDFAndreAinda não há avaliações
- Estudo Dirigido - Glicólise PDFDocumento2 páginasEstudo Dirigido - Glicólise PDFCatarina Rodrigues50% (2)
- Exercícios Bioquímica - RespostasDocumento29 páginasExercícios Bioquímica - RespostasThalita Yock88% (8)
- Material de Apoio para EstudoDocumento6 páginasMaterial de Apoio para EstudoNathália MeloAinda não há avaliações
- Fosforolização OxidativaDocumento16 páginasFosforolização Oxidativabrunowarley1982Ainda não há avaliações
- Fotossintese 3Documento13 páginasFotossintese 3Máicon KilppAinda não há avaliações
- Estudo Dirigido I UnidadeDocumento9 páginasEstudo Dirigido I UnidadeLUÍS FELIPE SANTANA AGUIAR100% (1)
- Estudo Dirigido - Metabolismo EnergeticoDocumento10 páginasEstudo Dirigido - Metabolismo EnergeticoVanessa Meirelles100% (2)
- AD2 Bioquímica IIDocumento2 páginasAD2 Bioquímica IIGabriel MagalhãesAinda não há avaliações
- Lista de Exercicios LipideosDocumento3 páginasLista de Exercicios LipideosMily SilvaAinda não há avaliações
- QB75E Lista Exerc 5Documento3 páginasQB75E Lista Exerc 5Lucineia Pinho0% (1)
- Introdução Ao MetabolismoDocumento38 páginasIntrodução Ao MetabolismoNayara GuedesAinda não há avaliações
- Estudo Dirigido BioquímicaDocumento9 páginasEstudo Dirigido BioquímicaMaria Eduarda BragaAinda não há avaliações
- Estudo DirigidoDocumento4 páginasEstudo DirigidoGabriel Moreira100% (1)
- Síntese ProteicaDocumento6 páginasSíntese ProteicaDííh GarciaAinda não há avaliações
- GlicoseDocumento8 páginasGlicoselaviAinda não há avaliações
- Ciclo de Krebs - Explicação QuímicaDocumento3 páginasCiclo de Krebs - Explicação QuímicaItamar Jesus100% (1)
- 1740718 (2).pptDocumento20 páginas1740718 (2).pptAle Araujo100% (1)
- Metabolismo CarboidratoDocumento9 páginasMetabolismo CarboidratoSabrina Xavier100% (1)
- Estudo Sobre Ácido Graxo - Lista ResolvidaDocumento8 páginasEstudo Sobre Ácido Graxo - Lista Resolvidajuliana100% (1)
- Bioquímica: Aula 3Documento16 páginasBioquímica: Aula 3Tuani de MirandaAinda não há avaliações
- Exercícios Introdução Ao MetabolismoDocumento13 páginasExercícios Introdução Ao MetabolismoanaAinda não há avaliações
- Cap. 8 - Metabolismo Energético Da CélulaDocumento12 páginasCap. 8 - Metabolismo Energético Da CélulaLuiz Pedro Lucas Da SilvaAinda não há avaliações
- Metabolismo Dos CarboidratosDocumento4 páginasMetabolismo Dos CarboidratosTaissa VitóriaAinda não há avaliações
- Metabolismo Dos CarboidratosDocumento4 páginasMetabolismo Dos CarboidratosTaissa VitóriaAinda não há avaliações
- Capítulo 3 - Estrutura e Funcionamento Da CélulaDocumento48 páginasCapítulo 3 - Estrutura e Funcionamento Da CélulaMegan Katherine Melling100% (2)
- Cola P2Documento3 páginasCola P2zepaflaAinda não há avaliações
- Metabolismo Dos Carboidratos e LipídiosDocumento4 páginasMetabolismo Dos Carboidratos e LipídiosNetto LacerdaAinda não há avaliações
- Metabolismo de Lipídeos KahioDocumento16 páginasMetabolismo de Lipídeos Kahiokahionz100% (2)
- Bioquimica - Digestão e Absorção Dos LipídiosDocumento5 páginasBioquimica - Digestão e Absorção Dos LipídiosRafael Rudá Coelho100% (1)
- Microsoft Word - Estudo Dirigido - Bloco LipídiosDocumento4 páginasMicrosoft Word - Estudo Dirigido - Bloco LipídiosFelipe MagalhaesAinda não há avaliações
- Aula 9 Metabolismo Dos LipídiosDocumento44 páginasAula 9 Metabolismo Dos Lipídiosaracnomarques100% (2)
- Aula 7-Digestão e Transporte de Lipídeos - 2022Documento59 páginasAula 7-Digestão e Transporte de Lipídeos - 2022Marco Antonio Damaceno100% (1)
- Ciclo de KrebsDocumento6 páginasCiclo de KrebsRichard WooliteAinda não há avaliações
- GlicóliseDocumento3 páginasGlicóliseTay100% (1)
- Metabolismo MicrobianoDocumento5 páginasMetabolismo MicrobianokassoroAinda não há avaliações
- Bioquímica - Aula 3 - CarboidratosDocumento38 páginasBioquímica - Aula 3 - CarboidratosElitePoa RsAinda não há avaliações
- Relatório 2 - Bioquímica 2 - Dosagem de Glicose e Fermentação em Saccharomyces CerevisiaeDocumento22 páginasRelatório 2 - Bioquímica 2 - Dosagem de Glicose e Fermentação em Saccharomyces CerevisiaeJheyce Moraes0% (1)
- Resumos de BioenergéticaDocumento22 páginasResumos de BioenergéticaHugo NicolauAinda não há avaliações
- 2 Prova de BioquímicaDocumento6 páginas2 Prova de BioquímicaMary FreitasAinda não há avaliações
- Enzimas (Questões Respondidas)Documento3 páginasEnzimas (Questões Respondidas)Nayara SantiagoAinda não há avaliações
- GlicóliseDocumento1 páginaGlicóliseConsolia DibAinda não há avaliações
- P1 Bioquímica PROVA 1Documento6 páginasP1 Bioquímica PROVA 1jaraAinda não há avaliações
- Lista Exercícios 2 - Bioquímica MetabólicaDocumento1 páginaLista Exercícios 2 - Bioquímica MetabólicaLucas PintanelAinda não há avaliações
- Lista de Exercícios - Bioquimica - 1 PDFDocumento4 páginasLista de Exercícios - Bioquimica - 1 PDFFabio Cavalcante da Silva0% (1)
- PEROXISSOMOSDocumento4 páginasPEROXISSOMOSCíntia FariaAinda não há avaliações
- Questões de BioquímicaDocumento8 páginasQuestões de BioquímicavalentinAinda não há avaliações
- Roteiro 1 - Metabolismo de Carboidratos!Documento3 páginasRoteiro 1 - Metabolismo de Carboidratos!Mariana ArgenteAinda não há avaliações
- Exercicos AminoácidosDocumento10 páginasExercicos AminoácidospscacaoAinda não há avaliações
- Fisiologia Hepática PDFDocumento5 páginasFisiologia Hepática PDFRenata Lima de FreitasAinda não há avaliações
- Gabarito de Biologia Celular - Prova 01Documento3 páginasGabarito de Biologia Celular - Prova 01Thaís LuzAinda não há avaliações
- Prova Bioquimica AB 2 CompletaDocumento3 páginasProva Bioquimica AB 2 CompletaEversonAinda não há avaliações
- Metabolismo de LipídiosDocumento26 páginasMetabolismo de LipídiosAmanda Luíze100% (3)
- BioenergéticaDocumento44 páginasBioenergéticaFranciedna AlcobaçaAinda não há avaliações
- Da Antiguidade à Redescoberta das Leis de MendelNo EverandDa Antiguidade à Redescoberta das Leis de MendelAinda não há avaliações
- FT 2-Ob Materia Autotroficos Sem CorrecaoDocumento4 páginasFT 2-Ob Materia Autotroficos Sem Correcaojoaomanuel monteiroteixeiraAinda não há avaliações
- Apostila Modulo XI Biossintese de LipideosDocumento33 páginasApostila Modulo XI Biossintese de LipideosVinicius RiannAinda não há avaliações
- Ae Bg10 B PPT FotossinteseDocumento16 páginasAe Bg10 B PPT FotossinteseMarcus LimaAinda não há avaliações
- Bqi 100 - Aula 08 - Catabolismo de CarboidratosDocumento33 páginasBqi 100 - Aula 08 - Catabolismo de CarboidratosAlícia TerbutinoAinda não há avaliações
- AutotrofiaDocumento23 páginasAutotrofiaLucas Pimentel AlemãoAinda não há avaliações
- Mapa MetabolicoDocumento1 páginaMapa MetabolicoAlisson MahechaAinda não há avaliações
- Metabolismo e Catabolismo - BioquimicaDocumento28 páginasMetabolismo e Catabolismo - BioquimicaHermes Sales CamposAinda não há avaliações
- Bio Ficha Unidade 2Documento17 páginasBio Ficha Unidade 2Matilde RodriguesAinda não há avaliações
- Aula 5 - Fotossíntese - Reações de CarboxilaçãoDocumento35 páginasAula 5 - Fotossíntese - Reações de CarboxilaçãoLuana Rodrigues CastiglioniAinda não há avaliações
- SimuladoDocumento7 páginasSimuladoManu DenckAinda não há avaliações
- Ficha Fotossintese VI CorreçãoDocumento1 páginaFicha Fotossintese VI CorreçãoVladyslav BaranenkoAinda não há avaliações
- Bioquímica MetabólicaDocumento3 páginasBioquímica MetabólicaGiovanna FerreiraAinda não há avaliações
- LipídiosDocumento25 páginasLipídiosMaria Rita SalesAinda não há avaliações
- Bioquímica 2 - Síntese de Acídos GraxosDocumento10 páginasBioquímica 2 - Síntese de Acídos GraxosGiulliene SousaAinda não há avaliações
- Metabolismo Do ColesterolDocumento54 páginasMetabolismo Do ColesterolAlvezz MarcoAinda não há avaliações
- Morfofisiologia Vegetal: Faculdade de Ciências BiológicasDocumento60 páginasMorfofisiologia Vegetal: Faculdade de Ciências BiológicasgonjurAinda não há avaliações
- Ficha Transportes FotossinteseDocumento6 páginasFicha Transportes Fotossintesegranja2023Ainda não há avaliações
- 8Bioq2L - Biossintese de Acidos Gordos - 2023s2 - 231110 - 120207Documento56 páginas8Bioq2L - Biossintese de Acidos Gordos - 2023s2 - 231110 - 120207Emerson De AguiarAinda não há avaliações
- Etanol Lignocelulosicorg PDFDocumento102 páginasEtanol Lignocelulosicorg PDFRanteAinda não há avaliações
- Aula10abril - Discussao - Stojkov22 - NETs - Fisiologia - Fisiopatologia - Viasmetabolicas (1) EM PORTUGUESDocumento21 páginasAula10abril - Discussao - Stojkov22 - NETs - Fisiologia - Fisiopatologia - Viasmetabolicas (1) EM PORTUGUESJEFFERSON VINICIUS DA SILVAAinda não há avaliações
- NAD+ No Envelhecimento Mecanismos Moleculares eDocumento5 páginasNAD+ No Envelhecimento Mecanismos Moleculares eRodrigo AlvesAinda não há avaliações
- Prova - Fisiologia VegetalDocumento23 páginasProva - Fisiologia VegetalvanderlanAinda não há avaliações
- FTRevisões CC FotossínteseDocumento12 páginasFTRevisões CC FotossíntesePaula LopesAinda não há avaliações
- Fotossintese 2014Documento113 páginasFotossintese 2014Josenildo Alves da SilvaAinda não há avaliações
- Apostila - Aula 1Documento53 páginasApostila - Aula 1rayane thaynara santosAinda não há avaliações