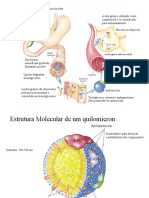
Você também pode gostar
- Metabolismo de LipídiosDocumento12 páginasMetabolismo de LipídiosDenisar PalmitoAinda não há avaliações
- Lista Oxidação de Ácidos GraxosDocumento6 páginasLista Oxidação de Ácidos GraxosGuilherme Belak SchmitkeAinda não há avaliações
- Oxidação 3 e 4 NovoDocumento2 páginasOxidação 3 e 4 NovoGuilherme Belak SchmitkeAinda não há avaliações
- Oxidação Dos Ácidos GraxosDocumento5 páginasOxidação Dos Ácidos GraxosBruna NevesAinda não há avaliações
- Metabolismo de LipídiosDocumento16 páginasMetabolismo de LipídiosDanielle RodriguesAinda não há avaliações
- Digestão, absorção e transporte de lipídiosDocumento11 páginasDigestão, absorção e transporte de lipídiosRicardo Junior100% (1)
- Capítulo 17Documento8 páginasCapítulo 17Davi SousaAinda não há avaliações
- Absorção de lipídeos (resumo Bia Mello)Documento2 páginasAbsorção de lipídeos (resumo Bia Mello)iagorochaAinda não há avaliações
- Bioquimica Digestao e Absorcao Dos LipidiosDocumento5 páginasBioquimica Digestao e Absorcao Dos LipidiosdalilaAinda não há avaliações
- 3.10.COC - Metabolismo 2Documento11 páginas3.10.COC - Metabolismo 2Christiane BarbiAinda não há avaliações
- Microsoft Word - Estudo Dirigido - Bloco LipídiosDocumento4 páginasMicrosoft Word - Estudo Dirigido - Bloco LipídiosFelipe MagalhaesAinda não há avaliações
- Oxidação dos ácidos graxos: via central de geração de energiaDocumento7 páginasOxidação dos ácidos graxos: via central de geração de energiaDariê CruvinelAinda não há avaliações
- Lipídios: funções, classificação e metabolismoDocumento82 páginasLipídios: funções, classificação e metabolismokaline cardosoAinda não há avaliações
- Aula 9 Metabolismo Dos LipídiosDocumento44 páginasAula 9 Metabolismo Dos Lipídiosaracnomarques100% (2)
- Catabolismo de Ácidos GraxosDocumento8 páginasCatabolismo de Ácidos Graxosjoão vitor fioratti de oliveiraAinda não há avaliações
- Metabolismo Do Colesterol - ResumidoDocumento5 páginasMetabolismo Do Colesterol - ResumidoDianaAinda não há avaliações
- Bioquimica - Digestão e Absorção Dos LipídiosDocumento5 páginasBioquimica - Digestão e Absorção Dos LipídiosRafael Rudá Coelho100% (1)
- Metabolismo de Carboidratos - Resumo de EscritoDocumento14 páginasMetabolismo de Carboidratos - Resumo de EscritoGualberto Júnior100% (1)
- Sessão 4Documento8 páginasSessão 4Gabriel PiresAinda não há avaliações
- LÍPIDOS Metabolismo 11Documento60 páginasLÍPIDOS Metabolismo 11Mirian NaimaAinda não há avaliações
- Resolução Lista BioquimicaDocumento16 páginasResolução Lista BioquimicaVitor SouzaAinda não há avaliações
- Metabolismo dos carboidratos e lipídiosDocumento4 páginasMetabolismo dos carboidratos e lipídiosNetto LacerdaAinda não há avaliações
- Metabolismo Dos Carboidratos, Proteínas E GordurasDocumento3 páginasMetabolismo Dos Carboidratos, Proteínas E GordurasGustavo JesusAinda não há avaliações
- Destino dos macronutrientes após refeiçãoDocumento14 páginasDestino dos macronutrientes após refeiçãoGustavo Henrique SilvaAinda não há avaliações
- Metabolismo Energético Nos AnimaisDocumento19 páginasMetabolismo Energético Nos AnimaisPaulo HenriqueAinda não há avaliações
- Bioquímica 2 - Metabolismo Dos TriacilgliceróisDocumento7 páginasBioquímica 2 - Metabolismo Dos TriacilgliceróisGiulliene SousaAinda não há avaliações
- 005 Metabolismo de Lipídios (Catabolismo)Documento10 páginas005 Metabolismo de Lipídios (Catabolismo)Izabella Trapia0% (1)
- Metabolismo dos LipídeosDocumento68 páginasMetabolismo dos LipídeosLetícia CabralAinda não há avaliações
- Metabolismo de LipoproteínasDocumento2 páginasMetabolismo de LipoproteínasTatiane Romero100% (2)
- ED Lipogênse, Lipólise, Gliconeogênese e CetogêneseDocumento14 páginasED Lipogênse, Lipólise, Gliconeogênese e CetogêneseBianca Veiga LopesAinda não há avaliações
- Lipólise - Degradação dos Ácidos GraxosDocumento6 páginasLipólise - Degradação dos Ácidos GraxosIvone Pires PortoAinda não há avaliações
- Metabolismo de LipídeosDocumento5 páginasMetabolismo de LipídeosMarcela Lima de BritoAinda não há avaliações
- Metabolismo Dos Lipideos Da DietaDocumento4 páginasMetabolismo Dos Lipideos Da Dietacasefa1100% (1)
- Catabolismo de LipídiosDocumento10 páginasCatabolismo de LipídiosIhury JhonsonAinda não há avaliações
- No sistema digestório - Metabolismo dos LipidiosDocumento6 páginasNo sistema digestório - Metabolismo dos Lipidiosedson sordaneAinda não há avaliações
- Metabolismo Dos LipídiosDocumento5 páginasMetabolismo Dos LipídiosMariana BarbosaAinda não há avaliações
- 1740718 (2).pptDocumento20 páginas1740718 (2).pptAle Araujo100% (1)
- Bioquimica Prova 3Documento7 páginasBioquimica Prova 3LuanaAinda não há avaliações
- Metabolismo Dos TriacilgliceroisDocumento32 páginasMetabolismo Dos Triacilgliceroismorena222Ainda não há avaliações
- Aula 8 - Metabolismo Dos LipídeosDocumento61 páginasAula 8 - Metabolismo Dos LipídeosSandy RodriguesAinda não há avaliações
- Bioenergética citoplasmáticaDocumento8 páginasBioenergética citoplasmáticaLone GhostAinda não há avaliações
- Integração Do Metabolismo IntermediárioDocumento27 páginasIntegração Do Metabolismo IntermediárioMatheus Borges100% (1)
- Metabolismo de Lipídeos IQSC-USPDocumento53 páginasMetabolismo de Lipídeos IQSC-USPEvany Priscila100% (1)
- Metabolismo de lipídeos: β-oxidação, corpos cetônicos e sínteseDocumento20 páginasMetabolismo de lipídeos: β-oxidação, corpos cetônicos e sínteseVanderval SIlva de OliveiraAinda não há avaliações
- GlicídiosDocumento7 páginasGlicídiosnykolle ribeiroAinda não há avaliações
- Bioquímica - Catabolismo de AminoácidosDocumento7 páginasBioquímica - Catabolismo de AminoácidosMarcela Lima de BritoAinda não há avaliações
- Síntese de lipídeos e esteróidesDocumento3 páginasSíntese de lipídeos e esteróidesMary FreitasAinda não há avaliações
- Lipideos 3Documento42 páginasLipideos 3Diéssica AlvesAinda não há avaliações
- Transcrição Dos Áudios 2 Prova de Bioquímica Clínica IDocumento16 páginasTranscrição Dos Áudios 2 Prova de Bioquímica Clínica ICarol Fernandes de Assis100% (1)
- Lipogênese: como o excesso de carboidratos é armazenado como gorduraDocumento2 páginasLipogênese: como o excesso de carboidratos é armazenado como gorduraMarina De Souza Bastos100% (1)
- β-oxidação de lipídiosDocumento44 páginasβ-oxidação de lipídiosMaria Eduarda Frota100% (1)
- Metabolismo de lipídios e aminoácidos e integração metabólicaDocumento72 páginasMetabolismo de lipídios e aminoácidos e integração metabólicaAdriano MateusAinda não há avaliações
- Tia JocaDocumento11 páginasTia JocaThiago R. N. SilvaAinda não há avaliações
- 5 Metabolismo de Lipídios e Aminoácidos e Integração MetabólicaDocumento66 páginas5 Metabolismo de Lipídios e Aminoácidos e Integração MetabólicarilixgsimululadorAinda não há avaliações
- Bioquímica do fígado: funções, metabolismo de carboidratos, lipoproteínas e xenobióticosDocumento5 páginasBioquímica do fígado: funções, metabolismo de carboidratos, lipoproteínas e xenobióticosRafaela MendesAinda não há avaliações
- RevisãoDocumento6 páginasRevisãoe236610Ainda não há avaliações
- Metabolismo Do Lipídeos 6.3Documento2 páginasMetabolismo Do Lipídeos 6.3Juliany Paz SilvaAinda não há avaliações
- Aula Recursos Ergogênicos (Suplementação)Documento20 páginasAula Recursos Ergogênicos (Suplementação)Thiago Oliveira de SousaAinda não há avaliações
- Processamento de Carnes - UNIDADE II - Valor Nutricional Da Carne - Profa. Dra. Ariana PassosDocumento32 páginasProcessamento de Carnes - UNIDADE II - Valor Nutricional Da Carne - Profa. Dra. Ariana Passosantonia.passosAinda não há avaliações
- Nomenclatura Dos AcilglicerideosDocumento3 páginasNomenclatura Dos AcilglicerideosChano FelicianoAinda não há avaliações
- Bioquimica 2013 Acidos NucleicosDocumento130 páginasBioquimica 2013 Acidos NucleicosAtila LourenaAinda não há avaliações
- Aula 4 - Ciclo Da UreiaDocumento22 páginasAula 4 - Ciclo Da UreiaBruno Moreira Dos SantosAinda não há avaliações
- Organizacao Molecular Da CelulaDocumento15 páginasOrganizacao Molecular Da CelulaNizzy BacanizzyAinda não há avaliações
- Bioquímica Da CélulaDocumento18 páginasBioquímica Da CélulaAyme NascimentoAinda não há avaliações
- Lavizoo Muda - Bula - 20220Documento2 páginasLavizoo Muda - Bula - 20220Edson França RodriguesAinda não há avaliações
- Keto Diet Plant Based1Documento14 páginasKeto Diet Plant Based1Mayara HamúAinda não há avaliações
- Análises Físico-Químicas de AlimentosDocumento196 páginasAnálises Físico-Químicas de AlimentosRafaela AcostaAinda não há avaliações
- Sintese Do LeiteDocumento47 páginasSintese Do LeitenhairaAinda não há avaliações
- Biossíntese de LipídeosDocumento8 páginasBiossíntese de LipídeosIhury Jhonson100% (1)
- Planilha de Dieta Automática - O EctomonstroDocumento6 páginasPlanilha de Dieta Automática - O EctomonstroMax Lopes dos SantosAinda não há avaliações
- 01 - Introdução À Bioquímica - Aula de Revisão e Nivelamento de Conceitos BásicosDocumento63 páginas01 - Introdução À Bioquímica - Aula de Revisão e Nivelamento de Conceitos BásicosJoelia maiaAinda não há avaliações
- Atividade Bioquimica CelularDocumento7 páginasAtividade Bioquimica Celularninja0pudimAinda não há avaliações
- Aula - DislipidemiaDocumento22 páginasAula - DislipidemiaallansalvadorpereiraAinda não há avaliações
- Bases Macromeleculares Da Constituição CelularDocumento37 páginasBases Macromeleculares Da Constituição CelularAndryah PaolaAinda não há avaliações
- E-Book Lipídios - NAPEDocumento26 páginasE-Book Lipídios - NAPEDiego SantosAinda não há avaliações
- VitaminasDocumento2 páginasVitaminasVivian RodriguesAinda não há avaliações
- Aminoácidos, peptídeos e proteínas: estruturas e funçõesDocumento45 páginasAminoácidos, peptídeos e proteínas: estruturas e funçõesLucas BatistaAinda não há avaliações
- Tabela SupermecadoDocumento9 páginasTabela Supermecadoflaperreira02Ainda não há avaliações
- Listado Tecnologias NoPOSDocumento545 páginasListado Tecnologias NoPOSYOLANDA DIAZAinda não há avaliações
- Vitaminas e avitaminoses no ensino médioDocumento4 páginasVitaminas e avitaminoses no ensino médioYasmim FoisAinda não há avaliações
- Atividade Sobre Replicação, Transcricao e Traducao 2023Documento2 páginasAtividade Sobre Replicação, Transcricao e Traducao 2023João VictorAinda não há avaliações
- Catalogo NewMillen 2020 WhatsappDocumento52 páginasCatalogo NewMillen 2020 WhatsappAllan PachecoAinda não há avaliações
- Resolução Lista BioquimicaDocumento16 páginasResolução Lista BioquimicaVitor SouzaAinda não há avaliações
- Modulação Intestinal Murilo PereiraDocumento5 páginasModulação Intestinal Murilo PereiraFernanda Rocha100% (1)
- CETO tabela suplementosDocumento4 páginasCETO tabela suplementosNara GonçalvesAinda não há avaliações
- Mec - Aminoácidos e ProteínasDocumento29 páginasMec - Aminoácidos e ProteínasBelHell PlacasAinda não há avaliações
- Nutrição de codornas cria e recriaDocumento6 páginasNutrição de codornas cria e recriaalcantara10Ainda não há avaliações