Você também pode gostar
- Resumo Da Terceira Semana Do Desenvolvimento EmbrionárioDocumento2 páginasResumo Da Terceira Semana Do Desenvolvimento EmbrionárioClaudia Nunes100% (1)
- Tecidos Do Corpo e Tecidos VegetaisDocumento2 páginasTecidos Do Corpo e Tecidos VegetaisartbiolifeAinda não há avaliações
- 6 - CambioDocumento1 página6 - CambiothailansaAinda não há avaliações
- 6 - Apostila CâmbioDocumento10 páginas6 - Apostila CâmbioBarbosa Marcelo AndradeAinda não há avaliações
- For Fun 1Documento9 páginasFor Fun 1Bruna AlonsoAinda não há avaliações
- Resumo 8Documento2 páginasResumo 8Luis Paulo MeurerAinda não há avaliações
- Botânica GeralDocumento11 páginasBotânica GeralAba lichakaAinda não há avaliações
- Anatomia de CauleDocumento41 páginasAnatomia de CauleJéssica N. CostaAinda não há avaliações
- Manual de Botanica AnatomicaDocumento171 páginasManual de Botanica AnatomicaBetho Monthenegro100% (1)
- Biologia 10 Unidade 6Documento8 páginasBiologia 10 Unidade 6Sofia Coelho100% (1)
- Resumo Caps 22 e 23 Do RavenDocumento6 páginasResumo Caps 22 e 23 Do RavenAna Luisa Ibrahim BertolliAinda não há avaliações
- O Caule TrabalhoDocumento10 páginasO Caule TrabalhoNeideAinda não há avaliações
- Prática 4Documento8 páginasPrática 4BEATRIZ GOMES RAMOSAinda não há avaliações
- Tarefa - XilemaDocumento4 páginasTarefa - XilemaJoão Eduardo AzevedoAinda não há avaliações
- Meristema e Crescimento VegetalDocumento3 páginasMeristema e Crescimento VegetalAna ClaraAinda não há avaliações
- Continuacao Da Gastrulacao (Somitos, Celoma)Documento8 páginasContinuacao Da Gastrulacao (Somitos, Celoma)rosangelaAinda não há avaliações
- IFB - Embriologia IIDocumento10 páginasIFB - Embriologia IIEmanuel NatanaelAinda não há avaliações
- III Semana de DesenvolvimentoDocumento31 páginasIII Semana de DesenvolvimentoLarisse SalvadorAinda não há avaliações
- Botanica TrabalhoDocumento25 páginasBotanica TrabalhoJuliana PaixãoAinda não há avaliações
- Monografia CauleDocumento9 páginasMonografia Caulebruno_shigueruAinda não há avaliações
- Relatório Biologia - Transporte Nas Plantas PDFDocumento30 páginasRelatório Biologia - Transporte Nas Plantas PDFO Sérgio ConceiçãoAinda não há avaliações
- Resumo 9Documento2 páginasResumo 9Luis Paulo MeurerAinda não há avaliações
- Valencio W de BotanicaDocumento13 páginasValencio W de BotanicaMocheyAinda não há avaliações
- 4-2 Aula BiocelDocumento54 páginas4-2 Aula BiocelFelipeMarodinAinda não há avaliações
- Resumo 4Documento2 páginasResumo 4Luis Paulo MeurerAinda não há avaliações
- Biologia PPT - Botânica - Histologia VegetalDocumento19 páginasBiologia PPT - Botânica - Histologia VegetalBiologia ppt100% (2)
- Sistema VascularDocumento13 páginasSistema VascularJhefferson GleizerAinda não há avaliações
- Resumo - FloemaDocumento9 páginasResumo - FloemaOcileudo SantosAinda não há avaliações
- Primeira e Segunda Fase Do Desenvolvimento EmbrionárioDocumento2 páginasPrimeira e Segunda Fase Do Desenvolvimento EmbrionárioJulianaAinda não há avaliações
- Resumão Do Corujão - Embrio - Prova 2 - 1) Gastrulação (Aula 1)Documento6 páginasResumão Do Corujão - Embrio - Prova 2 - 1) Gastrulação (Aula 1)jairofioraAinda não há avaliações
- Crescimento Secundário de Raiz e CauleDocumento8 páginasCrescimento Secundário de Raiz e CaulePâmella GomesAinda não há avaliações
- Estudo Dirigido de EmbriologiaDocumento6 páginasEstudo Dirigido de EmbriologiaMariele Tesche KüsterAinda não há avaliações
- Sistema UrogenitalDocumento6 páginasSistema UrogenitalKarin TreichelAinda não há avaliações
- ReproduçãoDocumento7 páginasReproduçãoAndré DelgadoAinda não há avaliações
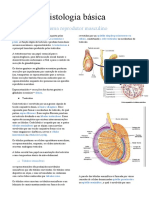
- Histologia Básic Sistema Reprodutor MasculinoDocumento5 páginasHistologia Básic Sistema Reprodutor MasculinoKlarissa LimaAinda não há avaliações
- Anatatomia Vegetal - TecidosDocumento5 páginasAnatatomia Vegetal - TecidosAvelino ChicoAinda não há avaliações
- Aula 3. Gastrulacao - 3 SemanaDocumento43 páginasAula 3. Gastrulacao - 3 SemanaAuryelle Sousa AzevedoAinda não há avaliações
- A Hematopoese - A Formação Do SangueDocumento10 páginasA Hematopoese - A Formação Do SangueCristiano Gley Carvalho MoreiraAinda não há avaliações
- Estudo Fisiologia VegetalDocumento4 páginasEstudo Fisiologia VegetalIsabellaAinda não há avaliações
- Botanica Geral Revisao 1 - Anatomia VegetalDocumento45 páginasBotanica Geral Revisao 1 - Anatomia VegetalbzintongaAinda não há avaliações
- Desenvolvimento Do Sistema CirculatórioDocumento5 páginasDesenvolvimento Do Sistema CirculatórioGabriel Ferro Roque100% (2)
- Fisiologia VegetalDocumento11 páginasFisiologia VegetalAílton GuambeAinda não há avaliações
- Revisão de Embriologia - 1 A 3 SemanaDocumento5 páginasRevisão de Embriologia - 1 A 3 SemanaleticiaAinda não há avaliações
- Condução Das SeivasDocumento8 páginasCondução Das SeivasEliseu Júlio MendesAinda não há avaliações
- Apostila Anatomia Vegetal UFUDocumento83 páginasApostila Anatomia Vegetal UFUKelly Belini0% (1)
- Divisao Da MitoseDocumento11 páginasDivisao Da MitoseAnonymous rgPNr8Xa7Ainda não há avaliações
- Trasnporte Nas Plantas e AnimaisDocumento7 páginasTrasnporte Nas Plantas e AnimaisMiriam FilipaAinda não há avaliações
- 3 Semana de DesenvolvimentoDocumento5 páginas3 Semana de DesenvolvimentoVicttoria Melo100% (1)
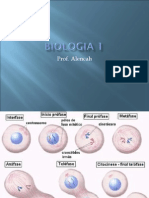
- Biologia PPT - Aula 14 Divisão CelularDocumento29 páginasBiologia PPT - Aula 14 Divisão CelularBiologia ppt100% (3)
- A Gastrulação É Um Estágio Importante Do Desenvolvimento EmbrionárioDocumento3 páginasA Gastrulação É Um Estágio Importante Do Desenvolvimento EmbrionárioAugusto UggioniAinda não há avaliações
- Xilema e Floema NaniDocumento30 páginasXilema e Floema NaniPablo Mendes CostaAinda não há avaliações
- Apostila Anatomia FloralDocumento13 páginasApostila Anatomia FloralIsabella CapillaAinda não há avaliações
- Histologia VegetalDocumento10 páginasHistologia VegetalLuisa MarquesAinda não há avaliações
- MeristemasDocumento1 páginaMeristemasschultzjeffersonAinda não há avaliações
- Resumo CitoesqueletoDocumento8 páginasResumo CitoesqueletoAdamastor NetoAinda não há avaliações
- ECTO MESO E ENDO Folheto Embrionário PDFDocumento8 páginasECTO MESO E ENDO Folheto Embrionário PDFfassisbr6137Ainda não há avaliações
- 9 - CauleDocumento1 página9 - CaulethailansaAinda não há avaliações
- Sistema de Transporte de Matéria Nas PlantasDocumento11 páginasSistema de Transporte de Matéria Nas PlantasJoão SilvaAinda não há avaliações
- 2aula Morfologia VegetalDocumento53 páginas2aula Morfologia VegetalLuiz Gustavo Peron MartinsAinda não há avaliações
- Resumo 9Documento2 páginasResumo 9Luis Paulo MeurerAinda não há avaliações
- Resumo 7Documento3 páginasResumo 7Luis Paulo MeurerAinda não há avaliações
- Resumo 10Documento2 páginasResumo 10Luis Paulo MeurerAinda não há avaliações
- Reumo 1Documento2 páginasReumo 1Luis Paulo MeurerAinda não há avaliações
- Resumo 5Documento2 páginasResumo 5Luis Paulo MeurerAinda não há avaliações
- Resumo 4Documento2 páginasResumo 4Luis Paulo MeurerAinda não há avaliações
- Resumo 3Documento3 páginasResumo 3Luis Paulo MeurerAinda não há avaliações
- Resumo 2Documento2 páginasResumo 2Luis Paulo MeurerAinda não há avaliações
- Relações Hidricas em Plantas - Aspectos AnatomicosDocumento16 páginasRelações Hidricas em Plantas - Aspectos AnatomicosAna Claudia MartinsAinda não há avaliações
- A FolhaDocumento6 páginasA FolhaParra1313Ainda não há avaliações
- Lista Histologia VegetalDocumento8 páginasLista Histologia Vegetalsamuel souzaAinda não há avaliações
- Fisiologia VegetalDocumento27 páginasFisiologia VegetalAriene Cristina Silva Prado100% (1)
- Trabalho Biologia Molecular Constituicao PlastideosDocumento11 páginasTrabalho Biologia Molecular Constituicao PlastideosFlorindoAinda não há avaliações
- Histologia Vegetal SlidesDocumento32 páginasHistologia Vegetal SlidesCor BanAinda não há avaliações
- ESTUDO DIRIGIDO-CascasDocumento7 páginasESTUDO DIRIGIDO-CascasIsabela HonoratoAinda não há avaliações
- Aula 3 - Tecidos Vegetais - Parte 1Documento29 páginasAula 3 - Tecidos Vegetais - Parte 1Carlos Martins100% (1)
- Aula Tropismos e NatismosDocumento26 páginasAula Tropismos e Natismosmirabilandia31Ainda não há avaliações
- 3 Meristemas Primários, Tecidos e Epiderme PDFDocumento30 páginas3 Meristemas Primários, Tecidos e Epiderme PDFMarcus Paulo FaustinoAinda não há avaliações
- Anatomia de RaizDocumento39 páginasAnatomia de RaizJéssica N. CostaAinda não há avaliações
- Os Anexos Da EpidermeDocumento7 páginasOs Anexos Da Epidermemilton shirakawaAinda não há avaliações
- Biologia - Fascículo 05 - Fisiologia VegetalDocumento10 páginasBiologia - Fascículo 05 - Fisiologia VegetalConcurso Vestibular100% (4)
- Tecidos VegetaisDocumento3 páginasTecidos VegetaisLeandro MontenegroAinda não há avaliações
- Relatório Nº 21Documento10 páginasRelatório Nº 21Joao FerreiraAinda não há avaliações
- XilemaDocumento1 páginaXilemavhuppes92Ainda não há avaliações
- BG10 27 Transporte Plantas 2021Documento50 páginasBG10 27 Transporte Plantas 2021Tiago CostaAinda não há avaliações
- Anatomia Dos Orgãos VegetaisDocumento31 páginasAnatomia Dos Orgãos VegetaisJunior BernardinoAinda não há avaliações
- Aula Prática 4 - MeristemasDocumento3 páginasAula Prática 4 - MeristemasReginaldo HenriqueAinda não há avaliações
- Prova II Diversidade de Fanerógamos UniasselviDocumento6 páginasProva II Diversidade de Fanerógamos UniasselviMauricio CamposAinda não há avaliações
- Germinação e Dormência de SementesDocumento7 páginasGerminação e Dormência de SementesDvrn DsgnAinda não há avaliações
- Fotossintese e RespiraçãoDocumento31 páginasFotossintese e RespiraçãoCharlesmaffraAinda não há avaliações
- Diversidade de FanerógamosDocumento8 páginasDiversidade de FanerógamosMauricio CamposAinda não há avaliações
- Estudo Dirigido Fisiologia VegetalDocumento7 páginasEstudo Dirigido Fisiologia Vegetaljosielly dantas de oliveiraAinda não há avaliações
- Texto de Apoio - Anatomia e Histologia VegetalDocumento27 páginasTexto de Apoio - Anatomia e Histologia VegetalANDRE SALVADOR MANJATEAinda não há avaliações
- Mapa Mental FitormoniosDocumento2 páginasMapa Mental FitormoniosOtavia Santos100% (1)
- Exercícios de Apoio - Transporte Nas PlantasDocumento8 páginasExercícios de Apoio - Transporte Nas PlantasRita SantosAinda não há avaliações
- Aula 4 Folha MorfologiaDocumento41 páginasAula 4 Folha MorfologiaJoão André Silveira da CostaAinda não há avaliações
- O Papel Dos Fitormônios No Desenvolvimento Das Plantas e Suas Utilidades EconômicasDocumento12 páginasO Papel Dos Fitormônios No Desenvolvimento Das Plantas e Suas Utilidades EconômicasDiego ArleyAinda não há avaliações