Você também pode gostar
- Defesa de Plantas Sinalizacao Quimica EEDocumento11 páginasDefesa de Plantas Sinalizacao Quimica EEEDUARDO OLIVEIRA GUILHERMEAinda não há avaliações
- Fitopatologia ADocumento113 páginasFitopatologia AVinícius Paim AlendeAinda não há avaliações
- Resumo Fatores de Virulência Nos FungosDocumento8 páginasResumo Fatores de Virulência Nos FungosRafaele Rodrigues100% (7)
- Sistema Imune de PlantasDocumento2 páginasSistema Imune de PlantasDenise Turini Gonzales Marioto0% (1)
- Comunicação Entre PlantasDocumento19 páginasComunicação Entre PlantasNádia OliveiraAinda não há avaliações
- Biotecnologia VegetalDocumento6 páginasBiotecnologia VegetalRodolfo Antônio da SilvaAinda não há avaliações
- Principiosepraticas RESISTENCIA DE INSETOS A INSETICIDASDocumento5 páginasPrincipiosepraticas RESISTENCIA DE INSETOS A INSETICIDASNádia Cristina OliveiraAinda não há avaliações
- Aula Fitopatologia 5 de AgostoDocumento44 páginasAula Fitopatologia 5 de AgostoAndre AlvesAinda não há avaliações
- Insetos SugadoresDocumento13 páginasInsetos SugadoresAN MarcosAinda não há avaliações
- Ed1 Imunologia ClinicaDocumento11 páginasEd1 Imunologia ClinicaEsthephani ClerAinda não há avaliações
- Interações Herbívoro - Planta e Suas Implicações para o Controle BiológicoDocumento21 páginasInterações Herbívoro - Planta e Suas Implicações para o Controle Biológicodarosa_fAinda não há avaliações
- RAPP 2014 Dalio Et Al - Final PDFDocumento40 páginasRAPP 2014 Dalio Et Al - Final PDFamandanacional1339Ainda não há avaliações
- Resistência de Pragas A PesticidasDocumento4 páginasResistência de Pragas A PesticidasDaniel TonettaAinda não há avaliações
- 1-s2 0-S0885576522000510-Main en PTDocumento9 páginas1-s2 0-S0885576522000510-Main en PTGenaina CristofoliAinda não há avaliações
- O Papel Do Sistema de Resistência Guarda Na Interação Gene-A-Gene em PlantasDocumento22 páginasO Papel Do Sistema de Resistência Guarda Na Interação Gene-A-Gene em PlantasRoberto Lanna FilhoAinda não há avaliações
- Modo de Ação TrichodermaDocumento4 páginasModo de Ação Trichodermaandressa.cazettaAinda não há avaliações
- Atividade - Resistência de Plantas - Plantas Inseticidas - Plantas TransgênicasDocumento2 páginasAtividade - Resistência de Plantas - Plantas Inseticidas - Plantas TransgênicasSâmilla TelesAinda não há avaliações
- 14 39 41 Apostilafitopatologia PDFDocumento60 páginas14 39 41 Apostilafitopatologia PDFIvaristo AmericoAinda não há avaliações
- Sebenta ImunologiaDocumento89 páginasSebenta ImunologiaRaquel MarquesAinda não há avaliações
- 16-Controle BiolgicoDocumento4 páginas16-Controle BiolgicoElias GomesAinda não há avaliações
- GD GeneticaDocumento5 páginasGD GeneticaAna Flávia BritoAinda não há avaliações
- GD GenéticaDocumento6 páginasGD Genéticagiovana nardiAinda não há avaliações
- 1991 Book MicropropagationDocumento11 páginas1991 Book Micropropagationhosana_rrAinda não há avaliações
- Fisiologia Do Parasitismo A Relacao Planta PatogenoDocumento16 páginasFisiologia Do Parasitismo A Relacao Planta PatogenoArlam SilvaAinda não há avaliações
- Resumo3 ECOPOP (P2) 2018Documento7 páginasResumo3 ECOPOP (P2) 2018Thais de Sousa SantosAinda não há avaliações
- Ecofisiologia VegetalDocumento6 páginasEcofisiologia VegetalmarinaAinda não há avaliações
- Agrotóxicos e DNADocumento8 páginasAgrotóxicos e DNAMichael MyersAinda não há avaliações
- DEfesa de PlantasDocumento18 páginasDEfesa de PlantasNádia OliveiraAinda não há avaliações
- Origem PragaDocumento13 páginasOrigem PragaDenio Carvalho MonteiroAinda não há avaliações
- Tipos de Fungicidas e DefiniçõesDocumento4 páginasTipos de Fungicidas e DefiniçõesJoshua ColemanAinda não há avaliações
- 1732 10898 1 PBDocumento8 páginas1732 10898 1 PBMárioHenriqueAinda não há avaliações
- Fungos Entomopatogênicos (Enzimas)Documento17 páginasFungos Entomopatogênicos (Enzimas)Juliana BarrosoAinda não há avaliações
- Imunidade Inata - Cap 2Documento13 páginasImunidade Inata - Cap 2Natália ChiesaAinda não há avaliações
- Metodos de Transformacao de PlantasDocumento15 páginasMetodos de Transformacao de PlantasGustavo PereiraAinda não há avaliações
- Controle Agroecológico de Pragas e DoençasDocumento4 páginasControle Agroecológico de Pragas e DoençasThaysa LimaAinda não há avaliações
- 02 - Interação Parasita-HospedeiroDocumento5 páginas02 - Interação Parasita-Hospedeiromaria eduardaAinda não há avaliações
- Fitopatologia Vegetal v.2Documento37 páginasFitopatologia Vegetal v.2Amanda NatháliaAinda não há avaliações
- Síndrome Pós Covid e MicrobiotaDocumento27 páginasSíndrome Pós Covid e MicrobiotaMoises AlbuquerqueAinda não há avaliações
- KALMUNEDocumento10 páginasKALMUNELetitia GonçalvesAinda não há avaliações
- Manual Fitossanidade PDFDocumento20 páginasManual Fitossanidade PDFAbílio Lima40% (5)
- Resumo ImunoDocumento17 páginasResumo ImunoBruna MartinsAinda não há avaliações
- 00 Controle de Pragas Insetos Rasteiros, Voadores - DedetizaçãoDocumento5 páginas00 Controle de Pragas Insetos Rasteiros, Voadores - DedetizaçãoAdriano Rosalim AdrilimAinda não há avaliações
- Plano de Aula - Inflamação X Infecção - Cintia Karina Elizandro e Cláudia Pacheco PedroDocumento15 páginasPlano de Aula - Inflamação X Infecção - Cintia Karina Elizandro e Cláudia Pacheco Pedrocintiake100% (1)
- Relatório 3Documento2 páginasRelatório 3Amanda BarbosaAinda não há avaliações
- Fitossanidade IntroduçãoDocumento17 páginasFitossanidade IntroduçãoCleiton DomingosAinda não há avaliações
- Métodos de Controle de PragasDocumento25 páginasMétodos de Controle de PragasMarcos Danilo Ramos mdr7Ainda não há avaliações
- Apostila Entomologia AgricolaDocumento310 páginasApostila Entomologia AgricolaAna Paula Saldanha100% (1)
- Do comensalismo à patogenicidade: Um estudo sobre estafilococos coagulase-negativaNo EverandDo comensalismo à patogenicidade: Um estudo sobre estafilococos coagulase-negativaAinda não há avaliações
- Microbiologia Médica I: Patógenos e Microbioma HumanoNo EverandMicrobiologia Médica I: Patógenos e Microbioma HumanoAinda não há avaliações
- Development of Eco-Friendly Agrochemicals a Modern Agricultural AlternativeNo EverandDevelopment of Eco-Friendly Agrochemicals a Modern Agricultural AlternativeAinda não há avaliações
- Fitorremediação em Solos Contaminados com HerbicidasNo EverandFitorremediação em Solos Contaminados com HerbicidasAinda não há avaliações
- Avaliação De Fungos Em Bioaerossóis Em Ambiente De Um Hospital De Médio Porte Do Noroeste PaulistaNo EverandAvaliação De Fungos Em Bioaerossóis Em Ambiente De Um Hospital De Médio Porte Do Noroeste PaulistaAinda não há avaliações
- Dieta GravidezDocumento2 páginasDieta GravidezCássioAdrianoAinda não há avaliações
- Folhas CandombleDocumento4 páginasFolhas Candombleapi-3730808100% (1)
- Tabelas CompletasDocumento9 páginasTabelas CompletasEdimar Lino NogueiraAinda não há avaliações
- Reprodução AssexuadaDocumento3 páginasReprodução AssexuadaitsbeeeaaaAinda não há avaliações
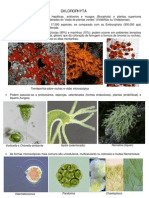
- ChlorophytaDocumento2 páginasChlorophytaRicardo JoppertAinda não há avaliações
- Questões de Raciocinio LogicoDocumento3 páginasQuestões de Raciocinio LogicoSPI CINTAinda não há avaliações
- Avaliação de Ciencias 4 BimesDocumento19 páginasAvaliação de Ciencias 4 BimesNilda Pereira LeiteAinda não há avaliações
- Dicionário Etimológico BiologiaDocumento2 páginasDicionário Etimológico BiologiaLucas VerasAinda não há avaliações
- Receitas Lanches SalgadosDocumento2 páginasReceitas Lanches SalgadosNicole Mantovani da SilvaAinda não há avaliações
- Manual de Despiese de LavadoraDocumento65 páginasManual de Despiese de LavadoraAdriana Muñoz MuñozAinda não há avaliações
- 11 - Ano - Classificação de WhittakerDocumento1 página11 - Ano - Classificação de WhittakerMárcio MonteiroAinda não há avaliações
- Apostila de Amala de SangoDocumento5 páginasApostila de Amala de SangoIfadareAinda não há avaliações
- Receita Indiana - Receitas VegetarianasDocumento7 páginasReceita Indiana - Receitas VegetarianasMaria Edith MarocaAinda não há avaliações
- FibonacciDocumento19 páginasFibonacciCristiano GomesAinda não há avaliações
- Horta em Mandala Adubada Com Microrganismos Eficientes, Com Consórcio de Plantas Repelentes para o Fortalecimento de BordadurasDocumento31 páginasHorta em Mandala Adubada Com Microrganismos Eficientes, Com Consórcio de Plantas Repelentes para o Fortalecimento de BordadurasGustavo SantanaAinda não há avaliações
- Dieta para DiabéticosDocumento18 páginasDieta para DiabéticosElmo Dutra Filho100% (1)
- Diagnose VisualDocumento26 páginasDiagnose VisualAndré FrançaAinda não há avaliações
- Magia Das ErvasDocumento13 páginasMagia Das ErvasChristine Ferreira Azzi71% (7)
- Efeitos Dos Agrotoxicos Sobre As Abelhas Silvestres BrasilDocumento88 páginasEfeitos Dos Agrotoxicos Sobre As Abelhas Silvestres BrasilRicardo Murilo Zanetti100% (1)
- Cartas BiologiaDocumento21 páginasCartas BiologiaAD Vilhena Congregação CanaãAinda não há avaliações
- Biologia - TaxonomiaDocumento41 páginasBiologia - TaxonomiaPablo MateusAinda não há avaliações
- Biologia Reprodutiva de Peixes OrnamentaisDocumento4 páginasBiologia Reprodutiva de Peixes OrnamentaishernanyAinda não há avaliações
- Agroindustria SemiaridoDocumento30 páginasAgroindustria SemiaridoAnny Kelly VasconcelosAinda não há avaliações
- Ficha de Avaliação de CN 8º Ano (Factores Abióticos e Bióticos Fluxo de Energia e Ciclo de Matéria)Documento4 páginasFicha de Avaliação de CN 8º Ano (Factores Abióticos e Bióticos Fluxo de Energia e Ciclo de Matéria)cristinapinheiro89% (9)
- GURPS - Grimório Verde - Biblioteca Élfica PDFDocumento29 páginasGURPS - Grimório Verde - Biblioteca Élfica PDFMárcio OliveiraAinda não há avaliações
- Prova Final EvaneideDocumento3 páginasProva Final EvaneideCarlos Luis Ribeiro Dos SantosAinda não há avaliações
- LichiaDocumento13 páginasLichiamorgxxxAinda não há avaliações
- CRUZADINHADocumento1 páginaCRUZADINHAgapasina88% (8)
- Guia de Problemas-1Documento47 páginasGuia de Problemas-1RenataAoun100% (1)
- 3.3.1. Caso 2 - Mr. Tangerine - Negociador 1Documento9 páginas3.3.1. Caso 2 - Mr. Tangerine - Negociador 1Zara TustraAinda não há avaliações