Você também pode gostar
- DownloadDocumento8 páginasDownloaddfssAinda não há avaliações
- Rejuvenescer Com O Plasma Sanguíneo Dos JovensNo EverandRejuvenescer Com O Plasma Sanguíneo Dos JovensAinda não há avaliações
- Recovery Nutry Tex-OeDocumento11 páginasRecovery Nutry Tex-OeMarcio CampiaoAinda não há avaliações
- HeiaDocumento1 páginaHeiaviviAinda não há avaliações
- Fisiologia Do Estresse e Sua InfluenciaDocumento13 páginasFisiologia Do Estresse e Sua InfluenciaEllen TelacioAinda não há avaliações
- Módulo 3 - Adaptações Celulares Ao EstresseDocumento33 páginasMódulo 3 - Adaptações Celulares Ao Estresseescola estadualAinda não há avaliações
- Resumo - Adaptação CelularDocumento4 páginasResumo - Adaptação CelularJoao Gabriel Chaves BequimamAinda não há avaliações
- Adaptação dos Ruminantes aos Climas QuentesNo EverandAdaptação dos Ruminantes aos Climas QuentesAinda não há avaliações
- Introdução À PatologiaDocumento23 páginasIntrodução À PatologiaAna Kelia Silva Melo100% (1)
- Algumas Considerações Sobre o Paciente Cirúrgico e A AnsiedadeDocumento6 páginasAlgumas Considerações Sobre o Paciente Cirúrgico e A AnsiedadePatrícia Rigoni DantasAinda não há avaliações
- Fisiologia e Fisiopatologia Do StressDocumento16 páginasFisiologia e Fisiopatologia Do StressPerisson DantasAinda não há avaliações
- Resumo - Lesões Celulares e Respostas Adaptativas PDFDocumento3 páginasResumo - Lesões Celulares e Respostas Adaptativas PDFDaniAinda não há avaliações
- Fisiologia Do EstresseDocumento27 páginasFisiologia Do EstressejoserobertopereiraAinda não há avaliações
- Estrutura Do MHC e Função - Apresentação de Antígenos. Parte 2Documento6 páginasEstrutura Do MHC e Função - Apresentação de Antígenos. Parte 2Erick GuerraAinda não há avaliações
- Relação Entre Apoptose e Alzheimer - BioquímicaDocumento13 páginasRelação Entre Apoptose e Alzheimer - BioquímicagleysonlopesAinda não há avaliações
- Fisiologia Do Estresse Aspectos Neuroendócrinos eDocumento11 páginasFisiologia Do Estresse Aspectos Neuroendócrinos eBruno Cerpa100% (1)
- Texto Características Dos Seres Vivos - Níveis de OrganizaçãoDocumento4 páginasTexto Características Dos Seres Vivos - Níveis de OrganizaçãoKelma Cristina de FreitasAinda não há avaliações
- Aspectos Fisiológicos Do EstresseDocumento4 páginasAspectos Fisiológicos Do EstressefranciscaAinda não há avaliações
- Tde 2Documento4 páginasTde 2Danizinha NünezAinda não há avaliações
- 3 - Reticulo Endoplasmatico - Controle de Qualidade Traducao PDFDocumento11 páginas3 - Reticulo Endoplasmatico - Controle de Qualidade Traducao PDFJoão Vitor Silva MendonçaAinda não há avaliações
- Apostila2012fisioi PDFDocumento118 páginasApostila2012fisioi PDFWeverton Jose NetoAinda não há avaliações
- 3361-Texto Do Artigo-10631-1-10-20181107Documento31 páginas3361-Texto Do Artigo-10631-1-10-20181107Rodrigo CampedelliAinda não há avaliações
- Apostila Bem - Estar e Producao AnimalDocumento29 páginasApostila Bem - Estar e Producao AnimalPaulo Afonso PaiãoAinda não há avaliações
- 01 - Lesão e Adaptação CelularDocumento26 páginas01 - Lesão e Adaptação CelularStanislav Ozzy Petrov100% (1)
- Livro Atualizado 1 PDFDocumento269 páginasLivro Atualizado 1 PDFMarlyne Ferreira PedrosaAinda não há avaliações
- 1 - Aula 2 - Mecan de Defesa - Adaptação CelularDocumento117 páginas1 - Aula 2 - Mecan de Defesa - Adaptação CelularJousneid BenilzaAinda não há avaliações
- Controle Da Expressão Gênica em Eucariotos-EpigenéticaDocumento4 páginasControle Da Expressão Gênica em Eucariotos-EpigenéticaSamanta Orana Pereira SampaioAinda não há avaliações
- 01 Introducao A BiologiaDocumento6 páginas01 Introducao A BiologiaSamuelSamuelAinda não há avaliações
- OTecido Adiposo Como Centro Reguladordo MetabolismoDocumento14 páginasOTecido Adiposo Como Centro Reguladordo Metabolismoalexsandra santosAinda não há avaliações
- 5632-Texto Do Artigo-18596-1-10-20221206Documento8 páginas5632-Texto Do Artigo-18596-1-10-20221206Gabriele CasagrandeAinda não há avaliações
- Estresse AnimalDocumento7 páginasEstresse AnimalEunice SantiagoAinda não há avaliações
- 01 Introducao A FisiologiaDocumento7 páginas01 Introducao A FisiologiaNicole Henriques Dos SantosAinda não há avaliações
- 844c84423cfcd7e05d2720770d2ee271Documento8 páginas844c84423cfcd7e05d2720770d2ee271gld silva100% (1)
- Influência Do Exercício Físico Na Resposta ImuneDocumento6 páginasInfluência Do Exercício Físico Na Resposta ImuneAlen_DAinda não há avaliações
- Sebenta Fisiologia HumanaDocumento92 páginasSebenta Fisiologia HumanaTania AlvesAinda não há avaliações
- O Ambiente, Função E Performance Animal: Rolim de Moura - Ro SETEMBRO/2023Documento7 páginasO Ambiente, Função E Performance Animal: Rolim de Moura - Ro SETEMBRO/2023Luiz Pedro Da VitoriaAinda não há avaliações
- Sp1.1 - Boff Oliveira, 2021Documento12 páginasSp1.1 - Boff Oliveira, 2021Tamires Nunes FariasAinda não há avaliações
- Células T Regulatórias Naturais (TREGS) em Doenças Reumáticas PDFDocumento14 páginasCélulas T Regulatórias Naturais (TREGS) em Doenças Reumáticas PDFRenatAvelarAinda não há avaliações
- Os Hormônios São Substâncias Químicas Mensageiras Produzidas Pelas GlândulasDocumento1 páginaOs Hormônios São Substâncias Químicas Mensageiras Produzidas Pelas GlândulasLara Silveira098Ainda não há avaliações
- DMSODocumento6 páginasDMSOAdriana GuimarãesAinda não há avaliações
- Fisiologia Veterinaria - ApostilaDocumento120 páginasFisiologia Veterinaria - ApostilaCaiane100% (2)
- Nihms 1527238Documento21 páginasNihms 1527238Makanudo.Ainda não há avaliações
- TMP 408 DDocumento13 páginasTMP 408 DFrontiersAinda não há avaliações
- RAPP 2014 Dalio Et Al - Final PDFDocumento40 páginasRAPP 2014 Dalio Et Al - Final PDFamandanacional1339Ainda não há avaliações
- 2019-Dissociação Adaptativa Entre Apoptose e NeurorregeneraçãoDocumento26 páginas2019-Dissociação Adaptativa Entre Apoptose e NeurorregeneraçãoEDEN GUADALUPE SIQUEIRAAinda não há avaliações
- Aula 01. PatologiaDocumento36 páginasAula 01. PatologiaBernardo gamesAinda não há avaliações
- Estresse Eixo HipofisarioDocumento49 páginasEstresse Eixo HipofisariossilveiracarlaAinda não há avaliações
- Avaliação 1 Matheus SantanaDocumento3 páginasAvaliação 1 Matheus SantanaMatheus SantanaAinda não há avaliações
- Fisiologia Veterinária IDocumento120 páginasFisiologia Veterinária Ihayane ramiresAinda não há avaliações
- O Estresse e As Doenças PsicossomáticasDocumento17 páginasO Estresse e As Doenças PsicossomáticasAntonio Fernando NavarroAinda não há avaliações
- Odis11 eNL Teste2Documento8 páginasOdis11 eNL Teste2santiagoformadorAinda não há avaliações
- Defesa de Plantas Sinalizacao Quimica EEDocumento11 páginasDefesa de Plantas Sinalizacao Quimica EEEDUARDO OLIVEIRA GUILHERMEAinda não há avaliações
- YamadaSouzaJuniorPereira TreinamentodeforaHipertrofiaeInflamao ArqMov2010Documento21 páginasYamadaSouzaJuniorPereira TreinamentodeforaHipertrofiaeInflamao ArqMov2010Muryllo Sirqueira Lopes Dos SantosAinda não há avaliações
- Fisiologia AnimalDocumento1 páginaFisiologia AnimalSimone CostaAinda não há avaliações
- Atividades Fisiologia - Conceitos Introdutórios Prof. Dr. Daniel Sol Sol de Medeiros HomeostasiaDocumento5 páginasAtividades Fisiologia - Conceitos Introdutórios Prof. Dr. Daniel Sol Sol de Medeiros HomeostasiaWilliam JorgeAinda não há avaliações
- Coração AnatomiaDocumento60 páginasCoração AnatomiaSónia RibeiroAinda não há avaliações
- Fisiologia Das Membranas FMUPDocumento22 páginasFisiologia Das Membranas FMUPSónia RibeiroAinda não há avaliações
- Titulacao Acido BaseDocumento7 páginasTitulacao Acido BaseSónia RibeiroAinda não há avaliações
- Torax, CV e Menbro SupDocumento12 páginasTorax, CV e Menbro SupSónia RibeiroAinda não há avaliações
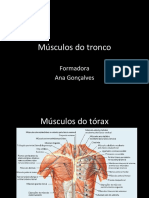
- Músculos Do TroncoDocumento70 páginasMúsculos Do TroncoSónia Ribeiro100% (1)
- CabecaDocumento12 páginasCabecaSónia RibeiroAinda não há avaliações
- GinecologiaDocumento549 páginasGinecologiaAntônio Kawakame NetoAinda não há avaliações
- Apostila Retentores PDFDocumento24 páginasApostila Retentores PDFLeandrodeLemos100% (1)
- Sortilégio Mistério Negro de ABDIAS DO NASCIMENTODocumento30 páginasSortilégio Mistério Negro de ABDIAS DO NASCIMENTOThiago EmanuelAinda não há avaliações
- Diagnóstico Dos Chakras Com Um PênduloDocumento5 páginasDiagnóstico Dos Chakras Com Um PênduloRonisson GuimaraesAinda não há avaliações
- Solucionário 4.2 PDFDocumento32 páginasSolucionário 4.2 PDFMatheus MonteiroAinda não há avaliações
- A Pura Verdade Sobre o NatalDocumento11 páginasA Pura Verdade Sobre o NatalfacescribdAinda não há avaliações
- 245-Texto Do Artigo-963-1-10-20161013Documento8 páginas245-Texto Do Artigo-963-1-10-20161013Bruno NunesAinda não há avaliações
- Resenha - A Arte Como LinguagemDocumento2 páginasResenha - A Arte Como LinguagemBernardo JunniorAinda não há avaliações
- Aula 2 - Material em SlideDocumento21 páginasAula 2 - Material em SlideAndré LuisAinda não há avaliações
- Diversidade LinguisticaDocumento22 páginasDiversidade LinguisticafernandesAinda não há avaliações
- Química Vestibulando DigitalDocumento216 páginasQuímica Vestibulando DigitalKleber Rogério100% (1)
- Exercícios - Passado Perfeito e ImperfeitoDocumento2 páginasExercícios - Passado Perfeito e ImperfeitoJoyceAinda não há avaliações
- 241 Lamina Qua e Qui 06 A 07 OutubrowebDocumento2 páginas241 Lamina Qua e Qui 06 A 07 OutubrowebJunior Baesso .Anaurilândia MSSAinda não há avaliações
- Apostila Catalise - 2014Documento195 páginasApostila Catalise - 2014Vitor RougemontAinda não há avaliações
- Estudo Do Concreto de Cimento Portland CDocumento61 páginasEstudo Do Concreto de Cimento Portland CMarcio ValerioAinda não há avaliações
- Zonas de Perigo Da FaceDocumento11 páginasZonas de Perigo Da Facemarcoslouro.medicinaAinda não há avaliações
- Exercícios de GeografiaDocumento21 páginasExercícios de GeografiaSandro Gomes0% (1)
- Atividade de Ciências 6º Ano (2)Documento5 páginasAtividade de Ciências 6º Ano (2)santosrochaariana2263100% (1)
- Utilização Do CimentoDocumento15 páginasUtilização Do CimentoRonivon OliveiraAinda não há avaliações
- Debret GEOGRAFIA PARTE ESCRITADocumento3 páginasDebret GEOGRAFIA PARTE ESCRITAGisele BelliniAinda não há avaliações
- MOD1 - UA1 - Resistência Dos Materiais II - 00 - 03Documento33 páginasMOD1 - UA1 - Resistência Dos Materiais II - 00 - 03dukathanAinda não há avaliações
- Classificação Periódica Dos ElementosDocumento19 páginasClassificação Periódica Dos ElementosSamuel SenaAinda não há avaliações
- Intermediário Ago - Ant FemininoDocumento4 páginasIntermediário Ago - Ant FemininoGabriel Bahia dos AnjosAinda não há avaliações
- Avaliação Diagnostica PortuguêsDocumento5 páginasAvaliação Diagnostica PortuguêsJoelma ResendeAinda não há avaliações
- Economia Exercicios de MicroeconomiaDocumento3 páginasEconomia Exercicios de MicroeconomiaportabilidadeamurelAinda não há avaliações
- NF Câmbio CVT Usado Netcar GasparDocumento1 páginaNF Câmbio CVT Usado Netcar GaspardiegoAinda não há avaliações
- Dds - Diálogo Diário de Saúde E SegurançaDocumento33 páginasDds - Diálogo Diário de Saúde E Segurançaalessandra.construtorarocaAinda não há avaliações
- Livro Física Das Radiações (Medicina Vibracional) (Carlos Assis)Documento163 páginasLivro Física Das Radiações (Medicina Vibracional) (Carlos Assis)CARLOS ASSISAinda não há avaliações
- Universidade Federal Da Bahia: Luiza BrandãoDocumento3 páginasUniversidade Federal Da Bahia: Luiza BrandãoLuíza BrandãoAinda não há avaliações
- Estruturas de MadeiraDocumento38 páginasEstruturas de Madeirajhonathann446Ainda não há avaliações
- Treinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNo EverandTreinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNota: 4.5 de 5 estrelas4.5/5 (169)
- Transtornos Alimentares: Entenda os aspectos que envolvem essas patologias e suas implicaçõesNo EverandTranstornos Alimentares: Entenda os aspectos que envolvem essas patologias e suas implicaçõesNota: 5 de 5 estrelas5/5 (2)
- RISCOS, VULNERABILIDADES E CONDICIONANTES URBANOSNo EverandRISCOS, VULNERABILIDADES E CONDICIONANTES URBANOSAinda não há avaliações
- Análise do Comportamento e suas Aplicações: Desafios e Possibilidades Volume 1No EverandAnálise do Comportamento e suas Aplicações: Desafios e Possibilidades Volume 1Nota: 5 de 5 estrelas5/5 (4)
- Uma voz sem palavras: Como o corpo libera o trauma e restaura o bem-estarNo EverandUma voz sem palavras: Como o corpo libera o trauma e restaura o bem-estarNota: 4.5 de 5 estrelas4.5/5 (9)
- Uma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNo EverandUma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNota: 4 de 5 estrelas4/5 (13)
- O Guia Da Apicultura Para Iniciantes: O Guia Completo Para Criar Abelhas, Aumentar Suas Colônias E Fazer Sua Colmeia ProsperarNo EverandO Guia Da Apicultura Para Iniciantes: O Guia Completo Para Criar Abelhas, Aumentar Suas Colônias E Fazer Sua Colmeia ProsperarAinda não há avaliações
- Biomecânica Básica dos Exercícios: Membros InferioresNo EverandBiomecânica Básica dos Exercícios: Membros InferioresNota: 3.5 de 5 estrelas3.5/5 (8)
- Fundamentos Em Cirurgia OralNo EverandFundamentos Em Cirurgia OralNota: 5 de 5 estrelas5/5 (1)
- Abismos Narcísicos: A Psicodinâmica do Amadurecimento e da IndividuaçãoNo EverandAbismos Narcísicos: A Psicodinâmica do Amadurecimento e da IndividuaçãoNota: 5 de 5 estrelas5/5 (2)
- Dieta pelo tipo sanguíneo: Da ancestralidade à epigenéticaNo EverandDieta pelo tipo sanguíneo: Da ancestralidade à epigenéticaNota: 5 de 5 estrelas5/5 (4)
- Bioquímica do corpo humano: As bases moleculares do metabolismoNo EverandBioquímica do corpo humano: As bases moleculares do metabolismoAinda não há avaliações