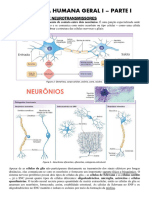
Você também pode gostar
- Capitulo 45 Resumo Fisiologia GuytonDocumento9 páginasCapitulo 45 Resumo Fisiologia Guytonmuller maiaAinda não há avaliações
- SinapsesDocumento13 páginasSinapsesNoemi SantosAinda não há avaliações
- Fisio Caso 1Documento21 páginasFisio Caso 1Julia JuliaAinda não há avaliações
- Estudo Dirigido de Membranas A NeurotransmissoresDocumento4 páginasEstudo Dirigido de Membranas A Neurotransmissoreskarina garzAinda não há avaliações
- Pricipais NeurotransmissoresDocumento9 páginasPricipais NeurotransmissoresJennifer Fontenelle100% (1)
- Exercicio Sinapse e NeurotransmissoresDocumento4 páginasExercicio Sinapse e NeurotransmissoresRaphael Gripp RibeiroAinda não há avaliações
- Sinápse PDFDocumento39 páginasSinápse PDFVandecir Silva100% (1)
- Pricipais NeurotransmissoresDocumento9 páginasPricipais NeurotransmissoresTarssila Rafaela Santos88% (8)
- Neurociência - 03 Mensageiros QuímicosDocumento2 páginasNeurociência - 03 Mensageiros Químicosbruno_marioto_1100% (1)
- Fisiologia Humana Geral I - Parte 1Documento39 páginasFisiologia Humana Geral I - Parte 1douglas.diasAinda não há avaliações
- Resumo Da Aula de NeurotransmissãoDocumento2 páginasResumo Da Aula de NeurotransmissãoRogério TavaresAinda não há avaliações
- Sinapses - FisiologiaDocumento4 páginasSinapses - FisiologiaLeonardo Moraes100% (1)
- Sinapse e NeurotransmissoresDocumento1 páginaSinapse e NeurotransmissoresJulia Cenci do CarmoAinda não há avaliações
- Fisiologia Humana 2Documento7 páginasFisiologia Humana 2Alessandro PedrettiAinda não há avaliações
- Neurofisiologia - Introdução À Neurotransmissão (Resumo)Documento3 páginasNeurofisiologia - Introdução À Neurotransmissão (Resumo)wiza934Ainda não há avaliações
- NeurotransmissoresDocumento16 páginasNeurotransmissoresFelipe Semente Lima100% (1)
- Bases NeurobiolôgicasDocumento3 páginasBases NeurobiolôgicasYrvynaAinda não há avaliações
- NeurotransmissoresDocumento11 páginasNeurotransmissoresLuciana Nogueira Coan100% (1)
- RESUMO - Fisiologia - Sistema NervosoDocumento14 páginasRESUMO - Fisiologia - Sistema NervosoIzaAinda não há avaliações
- Bioeletrogênese e Sinapses Dra. Ana Paula Peña DiasDocumento6 páginasBioeletrogênese e Sinapses Dra. Ana Paula Peña DiasarthurAinda não há avaliações
- Sinapse QuímicaDocumento23 páginasSinapse QuímicaErika Barreto100% (1)
- 4a Aula SinapseDocumento83 páginas4a Aula SinapseGabriel Messias100% (1)
- Sinapses e Junção NeuromuscularDocumento23 páginasSinapses e Junção NeuromuscularCamila MendesAinda não há avaliações
- Farmacologia II - 1 Etapa - DarlyDocumento88 páginasFarmacologia II - 1 Etapa - DarlyAna Luiza Sousa100% (1)
- NEURO - Encontros 4-5-6 PDFDocumento51 páginasNEURO - Encontros 4-5-6 PDFnaiara giovana gomes da silvaAinda não há avaliações
- Captação de Glicose Pelo MúsculoDocumento15 páginasCaptação de Glicose Pelo Músculovictorallan2005Ainda não há avaliações
- Bases Biológicas 3Documento11 páginasBases Biológicas 3victorallan2005Ainda não há avaliações
- Capítulo 8 SilverthornDocumento2 páginasCapítulo 8 SilverthornJean Carlo FlamiaAinda não há avaliações
- Neuro - Aula 4 - Sinapses e NeurotransmissoresDocumento33 páginasNeuro - Aula 4 - Sinapses e NeurotransmissoresMaíza Junior MartinsAinda não há avaliações
- Potencial de Ação e SinapsesDocumento9 páginasPotencial de Ação e SinapsesAmanda Francisquini100% (1)
- Estudo Dirigido 01 - Fisiologia - 2ºperíodoDocumento6 páginasEstudo Dirigido 01 - Fisiologia - 2ºperíodonetim69Ainda não há avaliações
- SinapseDocumento81 páginasSinapsecorretoraedisonreisimoveisAinda não há avaliações
- INTRODUÇÃO Ao SISTEMA NERVOSO - FisiologiaDocumento17 páginasINTRODUÇÃO Ao SISTEMA NERVOSO - Fisiologiacb_penatrujilloAinda não há avaliações
- Aula 04 Anatomia Fisiologia Da Sinapse PDFDocumento35 páginasAula 04 Anatomia Fisiologia Da Sinapse PDFGláucia Lima TocantynsAinda não há avaliações
- Neuroanatomia RESUMO TUDOOOOOOOODocumento36 páginasNeuroanatomia RESUMO TUDOOOOOOOOmelkinno50% (4)
- NEUROFISIOLOGIADocumento7 páginasNEUROFISIOLOGIAjoycebernardes2010Ainda não há avaliações
- Sinapse 1Documento75 páginasSinapse 1Luann Mikaell100% (1)
- Capítulo 16 - Resumo Biocel PDFDocumento10 páginasCapítulo 16 - Resumo Biocel PDFRapha BuenoAinda não há avaliações
- Mecanismos de Transdução de Sinal para Os Receptores de Membrana PlasmáticaDocumento12 páginasMecanismos de Transdução de Sinal para Os Receptores de Membrana PlasmáticaJacqueline Quintal100% (1)
- Bioquímica Metabolismo Do Sistema Nervoso 2Documento5 páginasBioquímica Metabolismo Do Sistema Nervoso 2aliciaabrantes26Ainda não há avaliações
- Mecanismo de SinapsesDocumento14 páginasMecanismo de SinapsesMakiesse António100% (1)
- Organização Do Sistema Nervoso Central, Funções Básicas Das Sinapses e NeurotransmissoresDocumento45 páginasOrganização Do Sistema Nervoso Central, Funções Básicas Das Sinapses e NeurotransmissoresLuciana OliveiraAinda não há avaliações
- Neurotransmissores e ReceptoreDocumento12 páginasNeurotransmissores e ReceptoreNara NeylaineAinda não há avaliações
- SinapseDocumento18 páginasSinapseJoão Ewerthon Barbosa LemosAinda não há avaliações
- Sistema Nervoso2Documento50 páginasSistema Nervoso2Anderson FreitasAinda não há avaliações
- Sistema NervosoDocumento27 páginasSistema Nervosoines santosAinda não há avaliações
- Fichamento - PsicofarmacologiaDocumento5 páginasFichamento - Psicofarmacologiapaulo.rpereiraAinda não há avaliações
- Biossíntese de NeurotransmissoresDocumento12 páginasBiossíntese de NeurotransmissoresIsadora RenóAinda não há avaliações
- Resumo Bixos Area 2 BioqDocumento35 páginasResumo Bixos Area 2 Bioqvinicius dinizAinda não há avaliações
- Receptores de Reserv 2Documento9 páginasReceptores de Reserv 2Larissa Acioli100% (1)
- (Condução Do Estímulo Nervoso 2014.2 ALUNOS) PDFDocumento21 páginas(Condução Do Estímulo Nervoso 2014.2 ALUNOS) PDFYasmin MattosAinda não há avaliações
- Aula 01 - COMUNICAÇÃO, SINALIZAÇÃO CELULAR E HOMEOSTASIADocumento7 páginasAula 01 - COMUNICAÇÃO, SINALIZAÇÃO CELULAR E HOMEOSTASIADaniel MunizAinda não há avaliações
- Resumo P1Documento24 páginasResumo P1LuanaAinda não há avaliações
- Processos de NeurotransmissãoDocumento8 páginasProcessos de NeurotransmissãoraianAinda não há avaliações
- Resumo Prova 01 Anestesiologia VeterináriaDocumento49 páginasResumo Prova 01 Anestesiologia VeterináriaBruno Félix MendesAinda não há avaliações
- Estudo Dirigido - P1 - Neurobiologia MolceularDocumento3 páginasEstudo Dirigido - P1 - Neurobiologia Molceulargabi.amota3Ainda não há avaliações
- Comunicação e Sinalização CelularDocumento14 páginasComunicação e Sinalização CelularGonçalo MachadoAinda não há avaliações
- Neurociências LivroDocumento4 páginasNeurociências LivroKarina LimaAinda não há avaliações
- Pontos Importantes de Cada TemaDocumento2 páginasPontos Importantes de Cada TemaRianne GomesAinda não há avaliações
- Resumo ReabilitaçãoDocumento8 páginasResumo ReabilitaçãoRianne GomesAinda não há avaliações
- Pena Pereira Torres, Eneida y Pereira (..) (2016) - Pesquisando Com o Metodo PsicanaliticoDocumento5 páginasPena Pereira Torres, Eneida y Pereira (..) (2016) - Pesquisando Com o Metodo PsicanaliticoBenicio :3Ainda não há avaliações
- Livro Psi Transito CRPDocumento129 páginasLivro Psi Transito CRPCarvalhoAinda não há avaliações
- Resumo Tema 5Documento6 páginasResumo Tema 5Rianne GomesAinda não há avaliações
- HistoriaDocumento7 páginasHistoriaWanderson ViltonAinda não há avaliações
- O Método Clínico e Sua Utilização Na PesquisaDocumento13 páginasO Método Clínico e Sua Utilização Na PesquisaRianne GomesAinda não há avaliações
- SILVA, C. M. MACEDO, M. M. K. O Método Psicanalítico de Pesquisa e A Potencialidade Dos Fatos ClínicosDocumento14 páginasSILVA, C. M. MACEDO, M. M. K. O Método Psicanalítico de Pesquisa e A Potencialidade Dos Fatos ClínicosRaphael AguiarAinda não há avaliações
- Appolinário, Fabio - Metodologia Da Ciência - Filosofia e Prática Da Pesquisa-Cengage Learning (2012)Documento240 páginasAppolinário, Fabio - Metodologia Da Ciência - Filosofia e Prática Da Pesquisa-Cengage Learning (2012)Kkgkkfmmm60% (5)
- 10 PDFDocumento10 páginas10 PDFAlexAinda não há avaliações
- 05Documento6 páginas05luizdilelio43Ainda não há avaliações
- Aplicações Do Método Fenomenológico À PesquisaDocumento10 páginasAplicações Do Método Fenomenológico À PesquisaGustavo SantosAinda não há avaliações
- O Rigor Da Pesquisa Psicanalítica Métodos de Avaliação de Sua Validade e ConfiabilidadeDocumento9 páginasO Rigor Da Pesquisa Psicanalítica Métodos de Avaliação de Sua Validade e ConfiabilidadeRianne GomesAinda não há avaliações
- Neuro - AlunosDocumento116 páginasNeuro - AlunosEllen AraujoAinda não há avaliações
- Prova de Fisiologia - EsamazDocumento5 páginasProva de Fisiologia - EsamazRafaela LobatoAinda não há avaliações
- UNIP Aps CertoDocumento16 páginasUNIP Aps CertoSuellen ReisAinda não há avaliações
- O Funcionamento Global Do Cérebro Humano - Especialização FuncionalDocumento3 páginasO Funcionamento Global Do Cérebro Humano - Especialização FuncionalMicael CiênciaAinda não há avaliações
- ED 2 - Farmacologia Aplicada Ao Sistema ColinérgicoDocumento6 páginasED 2 - Farmacologia Aplicada Ao Sistema ColinérgicoAdele LeiteAinda não há avaliações
- Diagnostico Por Ultrassom - Cabe - Anil AhujaDocumento1.405 páginasDiagnostico Por Ultrassom - Cabe - Anil AhujaElessandro FerreiraAinda não há avaliações
- Teoria Polivagal 04.04.23Documento34 páginasTeoria Polivagal 04.04.23Joao GuerreiroAinda não há avaliações
- Estudo Dirigido - Planos Eixos Esqueletico ArticularDocumento3 páginasEstudo Dirigido - Planos Eixos Esqueletico ArticularKeilliane NunesAinda não há avaliações
- 22) MetástasesDocumento60 páginas22) MetástasesAsdfAinda não há avaliações
- PORTF3Documento61 páginasPORTF3Nicole SantosAinda não há avaliações
- (Imagem) 3 - TomografiaDocumento5 páginas(Imagem) 3 - Tomografiavictor corrêaAinda não há avaliações
- Texto Base 02Documento5 páginasTexto Base 02EDSONAinda não há avaliações
- PatologiadosistemanervosoDocumento25 páginasPatologiadosistemanervosoProf. Margarida MachadoAinda não há avaliações
- Organização Celular Do Sistema NervosoDocumento3 páginasOrganização Celular Do Sistema NervosoGlória MamédioAinda não há avaliações
- Simulado 1 AnatomiaDocumento4 páginasSimulado 1 AnatomiaPaulo SouzaAinda não há avaliações
- Fisiologia Do Sistema RespiratórioDocumento21 páginasFisiologia Do Sistema RespiratórioAlícia HellenAinda não há avaliações
- Revisto - Órgãos e Visceras Revisado Set 12 2022 2Documento297 páginasRevisto - Órgãos e Visceras Revisado Set 12 2022 2Florentino CueAinda não há avaliações
- QUESTIONARIO Histologia OutubroDocumento5 páginasQUESTIONARIO Histologia OutubroKerly CarvalhoAinda não há avaliações
- 11 - Estrutura e Funções Do Tálamo, Subtálamo e EpitálamoDocumento4 páginas11 - Estrutura e Funções Do Tálamo, Subtálamo e EpitálamoReinaldo Riquet100% (1)
- Revisão Semiologia AV2Documento45 páginasRevisão Semiologia AV2Vanessa SilvérioAinda não há avaliações
- Apostila de Massoterapia AplicadaDocumento52 páginasApostila de Massoterapia AplicadaDimitry RodriguesAinda não há avaliações
- Capítulo S Limbico Fisiologia Aplicada À PsicologiaDocumento3 páginasCapítulo S Limbico Fisiologia Aplicada À PsicologiaValeskaSenaAinda não há avaliações
- TeoricoDocumento35 páginasTeoricoMariana AdeoAinda não há avaliações
- Introducao A HistologiaDocumento80 páginasIntroducao A HistologiaSarah De SousaAinda não há avaliações
- 2da2f1de Prova Residencia Medica Gabarito Sirio Libanes 2021Documento50 páginas2da2f1de Prova Residencia Medica Gabarito Sirio Libanes 2021KISCILLA BORGESAinda não há avaliações
- Slides MotilidadeDocumento19 páginasSlides Motilidademateus71314Ainda não há avaliações
- Roteiro de Neuromotricidade RespostasDocumento9 páginasRoteiro de Neuromotricidade RespostasBruno MoschiarAinda não há avaliações
- Anatomia Cruz VermelhaDocumento239 páginasAnatomia Cruz VermelhaLuís Filipe CostaAinda não há avaliações
- Imaginologia NeurológicaDocumento100 páginasImaginologia NeurológicaQuilza GomesAinda não há avaliações
- Aula 02 - Sistema Esquelético (Parte I) PDFDocumento77 páginasAula 02 - Sistema Esquelético (Parte I) PDFAlícia HellenAinda não há avaliações
- Equilíbrio iônico: aplicações em química analíticaNo EverandEquilíbrio iônico: aplicações em química analíticaAinda não há avaliações
- Química da Cerveja: Uma Abordagem Química e Bioquímica das Matérias-Primas, Processo de Produção e da Composição dos Compostos de Sabores da CervejaNo EverandQuímica da Cerveja: Uma Abordagem Química e Bioquímica das Matérias-Primas, Processo de Produção e da Composição dos Compostos de Sabores da CervejaAinda não há avaliações
- Série Didática para o Apoio a Formação de Professores de Química: Volume 2: MoléculasNo EverandSérie Didática para o Apoio a Formação de Professores de Química: Volume 2: MoléculasNota: 5 de 5 estrelas5/5 (1)
- Simetria Molecular III: Série Didática para o apoio a formação de professores de QuímicaNo EverandSimetria Molecular III: Série Didática para o apoio a formação de professores de QuímicaAinda não há avaliações
- Uma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNo EverandUma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNota: 4 de 5 estrelas4/5 (13)
- Química na Educação Básica: Ferramentas teóricas e práticasNo EverandQuímica na Educação Básica: Ferramentas teóricas e práticasNota: 4 de 5 estrelas4/5 (1)
- Uma breve história da química: Da alquimia às ciências moleculares modernasNo EverandUma breve história da química: Da alquimia às ciências moleculares modernasAinda não há avaliações
- Detetive Por Um Dia: Aprendendo A Investigar Por Meio Da Química ForenseNo EverandDetetive Por Um Dia: Aprendendo A Investigar Por Meio Da Química ForenseAinda não há avaliações
- Problemas de Cinética e Reatores Químicos: 100 Problemas Resolvidos, 500 Problemas Propostos (Com Respostas)No EverandProblemas de Cinética e Reatores Químicos: 100 Problemas Resolvidos, 500 Problemas Propostos (Com Respostas)Ainda não há avaliações
- Reologia do Concreto: Fatores influentes para previsão da retração e fluênciaNo EverandReologia do Concreto: Fatores influentes para previsão da retração e fluênciaAinda não há avaliações
- Química em tubos de ensaio: uma abordagem para principiantesNo EverandQuímica em tubos de ensaio: uma abordagem para principiantesAinda não há avaliações
- Vigilância Sanitária Nos Estados E MunicípiosNo EverandVigilância Sanitária Nos Estados E MunicípiosAinda não há avaliações
- Para Gostar de Química: Introdução à Química GeralNo EverandPara Gostar de Química: Introdução à Química GeralAinda não há avaliações
- Princípios de química analítica: Abordagem teórica qualitativa e quantitativaNo EverandPrincípios de química analítica: Abordagem teórica qualitativa e quantitativaNota: 5 de 5 estrelas5/5 (1)
- Uso do Policloreto de Alumínio (PAC): vantagens sobre o Sulfato de Alumínio em sistema de tratamento de águaNo EverandUso do Policloreto de Alumínio (PAC): vantagens sobre o Sulfato de Alumínio em sistema de tratamento de águaAinda não há avaliações
- Eletroanálises: aspectos teóricos e práticosNo EverandEletroanálises: aspectos teóricos e práticosAinda não há avaliações