Você também pode gostar
- Pneumonia em Pediatria - DiretrizesDocumento20 páginasPneumonia em Pediatria - Diretrizescason100% (1)
- Dossiê de Estágio de MicrobiologiaDocumento24 páginasDossiê de Estágio de MicrobiologiaMariana AnacletoAinda não há avaliações
- Riscos em EsteticaDocumento64 páginasRiscos em EsteticaRosângela VasconcelosAinda não há avaliações
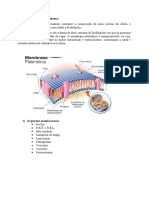
- UFRA Membrana plasmática funções permeabilidadeDocumento46 páginasUFRA Membrana plasmática funções permeabilidadequel04Ainda não há avaliações
- Ctic9 N6 Doenças e Saúde Do Sistema ReprodutorDocumento10 páginasCtic9 N6 Doenças e Saúde Do Sistema ReprodutorJoão Pedro Pinto SilvaAinda não há avaliações
- SISTEMA ABO - Prof. Adão Marcos Graciano Dos SantosDocumento3 páginasSISTEMA ABO - Prof. Adão Marcos Graciano Dos SantosAdão Marcos GracianoAinda não há avaliações
- Tipos de Mutações Genéticas e CromossómicasDocumento11 páginasTipos de Mutações Genéticas e CromossómicasLua NzingaAinda não há avaliações
- Apostila de Biologia - Vol 2Documento38 páginasApostila de Biologia - Vol 2Biologia Concurso Vestibular100% (2)
- Apostila de Biologia - Vol 2Documento38 páginasApostila de Biologia - Vol 2Biologia Concurso Vestibular100% (2)
- Resmo - Tráfego Intracelular de VesículasDocumento12 páginasResmo - Tráfego Intracelular de Vesículas9vvzhn8krvAinda não há avaliações
- Tráfego Intracelular de VesículasDocumento5 páginasTráfego Intracelular de VesículasjbeloniAinda não há avaliações
- Via secreção biologia celularDocumento35 páginasVia secreção biologia celularKarina Fernandes ValeAinda não há avaliações
- Tráfego VesicularDocumento15 páginasTráfego Vesicularanonimo anonimoAinda não há avaliações
- Aula 5 Tranporte VesicularDocumento36 páginasAula 5 Tranporte VesicularÉrica Boer GarciaAinda não há avaliações
- Estudo Dirigido - Compartimentos Intracelulares e Transporte de ProteínasDocumento2 páginasEstudo Dirigido - Compartimentos Intracelulares e Transporte de ProteínasLuiza LopesAinda não há avaliações
- Membrana Celular: Transporte e Estrutura emDocumento46 páginasMembrana Celular: Transporte e Estrutura emTatiane ValeAinda não há avaliações
- Transporte Intracitoplasmático de VesículasDocumento8 páginasTransporte Intracitoplasmático de VesículasLorena do Espirito Santo CostaAinda não há avaliações
- Membrana PlasmáticaDocumento10 páginasMembrana PlasmáticaNz NaganAinda não há avaliações
- Exercícios 1 CitologiaDocumento15 páginasExercícios 1 CitologiaRichard JuniorAinda não há avaliações
- Membrana Plasmática: Transporte e EstruturasDocumento32 páginasMembrana Plasmática: Transporte e EstruturasPriscila MarinAinda não há avaliações
- Resumo Biocel 1 Etapa - LuizaDocumento19 páginasResumo Biocel 1 Etapa - Luizaraf.marques.20Ainda não há avaliações
- Citologia fronteirasDocumento14 páginasCitologia fronteirasJuliana Luizetto de LuccaAinda não há avaliações
- Aula 4 Tranporte VesicularDocumento37 páginasAula 4 Tranporte VesicularAntonio Carlos Cruz Da SilvaAinda não há avaliações
- Estrutura e função das membranas celulares e sistemas de transporteDocumento14 páginasEstrutura e função das membranas celulares e sistemas de transporteLuis RicardoAinda não há avaliações
- Prova de Bio Cel ResumoDocumento8 páginasProva de Bio Cel ResumoThais GonzagaAinda não há avaliações
- Compartimentos Intracelulares e TransporteDocumento8 páginasCompartimentos Intracelulares e Transportefevabax924Ainda não há avaliações
- Exercicio Citologia RespondidoDocumento10 páginasExercicio Citologia RespondidoWanessa TeófilaAinda não há avaliações
- Estrutura das Membranas CelularesDocumento19 páginasEstrutura das Membranas CelularesThiago SantosAinda não há avaliações
- Células eucariontes: estrutura e funçãoDocumento5 páginasCélulas eucariontes: estrutura e funçãoLucas NevesAinda não há avaliações
- Membranas e transporte: tipos e mecanismosDocumento6 páginasMembranas e transporte: tipos e mecanismosSheila PinhoAinda não há avaliações
- Membrana CelularDocumento10 páginasMembrana CelularEsmeralda MuchisseAinda não há avaliações
- Biologia Celular e Genética - Aula 02 - Membrana Plasmática e Transporte de MembranaDocumento25 páginasBiologia Celular e Genética - Aula 02 - Membrana Plasmática e Transporte de MembranaDouglas NobreAinda não há avaliações
- Complexo de Golgi e LisossomosDocumento9 páginasComplexo de Golgi e LisossomosJúlia MottaAinda não há avaliações
- Resumo (Citologia)Documento3 páginasResumo (Citologia)Joana LobatoAinda não há avaliações
- A estrutura e função da membrana plasmáticaDocumento2 páginasA estrutura e função da membrana plasmáticaMaria Theresa MoreiraAinda não há avaliações
- Bio Aula Célula - Transporte - Divisão Celular - Embriologia - HistologiaDocumento7 páginasBio Aula Célula - Transporte - Divisão Celular - Embriologia - HistologiaMatheus PadilhaAinda não há avaliações
- Fechamento Problema 2Documento4 páginasFechamento Problema 2anna luisaAinda não há avaliações
- Sistema MembranarDocumento7 páginasSistema MembranarAlteza Pró100% (3)
- Compartimentos Intracelulares e Endereçamento de ProteínasDocumento11 páginasCompartimentos Intracelulares e Endereçamento de ProteínasGabriel Da Silva SengesAinda não há avaliações
- O transporte entre citosol e núcleoDocumento12 páginasO transporte entre citosol e núcleoMatheus Correa0% (1)
- Aula 3 2023Documento33 páginasAula 3 2023calmonhugo24Ainda não há avaliações
- Estudo Biologia2Documento9 páginasEstudo Biologia2WQWQWEQWEQAinda não há avaliações
- Aula MembranasDocumento33 páginasAula MembranasClaudio CirneAinda não há avaliações
- Estudo Dirigido - Minhas RespostasDocumento4 páginasEstudo Dirigido - Minhas RespostasNayara ChavesAinda não há avaliações
- Resumo da Homeostase Celular e Membrana PlasmáticaDocumento4 páginasResumo da Homeostase Celular e Membrana PlasmáticaDébora MartinsAinda não há avaliações
- Modulo 2 Introducao A Citologia e Membrana Plasmatica - Doc - 20240314 - 113055 - 0000Documento13 páginasModulo 2 Introducao A Citologia e Membrana Plasmatica - Doc - 20240314 - 113055 - 0000Akiane FelixAinda não há avaliações
- Membrana Plasmática e A PermeabilidadeDocumento39 páginasMembrana Plasmática e A PermeabilidadeWesleyJúniorAinda não há avaliações
- Biologia Molecular Da Célula (Alberts Et Al, 5 Ed.)Documento13 páginasBiologia Molecular Da Célula (Alberts Et Al, 5 Ed.)Rodrigo SimaoAinda não há avaliações
- Biofísica Das Membranas BiológicasDocumento23 páginasBiofísica Das Membranas BiológicasAlexandre Bastos Gomide50% (2)
- MI 1º ANO Unid 3 - Célula - Cap. 7a - Membrana PlasmáticaDocumento4 páginasMI 1º ANO Unid 3 - Célula - Cap. 7a - Membrana PlasmáticaEriczinAinda não há avaliações
- Complexo de Golgi: estrutura e funçõesDocumento5 páginasComplexo de Golgi: estrutura e funçõesForwomenAinda não há avaliações
- Transporte de Proteínas para Organelas por Sinais de DistribuiçãoDocumento5 páginasTransporte de Proteínas para Organelas por Sinais de DistribuiçãoMacklecio OliveiraAinda não há avaliações
- Membrana Plasmática - Características e Resumo - Mundo EducaçãoDocumento3 páginasMembrana Plasmática - Características e Resumo - Mundo EducaçãoMarcos Vinícius Neves OlsanAinda não há avaliações
- Membrana PlasmáticaDocumento6 páginasMembrana PlasmáticaLeticia KirchimairAinda não há avaliações
- 2986-2 - Citologia - Membrana PlasmáticaDocumento6 páginas2986-2 - Citologia - Membrana PlasmáticaRênisson KalinoAinda não há avaliações
- Estrutura da membrana21Documento6 páginasEstrutura da membrana21Monica AlexandreAinda não há avaliações
- CADERNO DE FISIOLOGIA - UFMA (2015.1).docxDocumento76 páginasCADERNO DE FISIOLOGIA - UFMA (2015.1).docxAmaury VieiraAinda não há avaliações
- Biofísica das MembranasDocumento23 páginasBiofísica das MembranasWagner PimentelAinda não há avaliações
- Biomembranas e transportes celularesDocumento47 páginasBiomembranas e transportes celularesAlex OjimaAinda não há avaliações
- Membrana PlasmáticaDocumento5 páginasMembrana PlasmáticaNayara SilvaAinda não há avaliações
- Histologia - CitologiaDocumento12 páginasHistologia - CitologiaPhelipe MonteiroAinda não há avaliações
- Funções organelas célula vegetal animalDocumento3 páginasFunções organelas célula vegetal animalThaís LuzAinda não há avaliações
- CEDERJ Biologia Celular I - Gabarito 13 A 20Documento9 páginasCEDERJ Biologia Celular I - Gabarito 13 A 20api-3800070Ainda não há avaliações
- Fisiopatologia Pancreas Exocrino-CoresDocumento11 páginasFisiopatologia Pancreas Exocrino-CoresCarla Isabel Pereira100% (1)
- Transmissão, regulação e operão da lactose em testes de biologiaDocumento3 páginasTransmissão, regulação e operão da lactose em testes de biologiaEduarda Rosa Vieira SantosAinda não há avaliações
- Alegrete Curso Inspeção Sanitária LaboratorioDocumento21 páginasAlegrete Curso Inspeção Sanitária LaboratorioBernadete CerattiAinda não há avaliações
- Anticorpos e imunidadeDocumento5 páginasAnticorpos e imunidadeJoyce FerreiraAinda não há avaliações
- Gerar Laudos PDFDocumento1 páginaGerar Laudos PDFNati BurlaccineAinda não há avaliações
- DNA para RNA e proteínasDocumento3 páginasDNA para RNA e proteínasSheila Santos0% (1)
- ATLAS Diagnostico Leishmaniose Tegumentar AmericanaDocumento138 páginasATLAS Diagnostico Leishmaniose Tegumentar AmericanaJuan Camilo Sánchez Arcila100% (2)
- A Molécula Da VidaDocumento10 páginasA Molécula Da VidaJUNIOR100% (1)
- Tabela SusDocumento5 páginasTabela SusCristineAinda não há avaliações
- Antivirais para o tratamento do HIVDocumento2 páginasAntivirais para o tratamento do HIVKlecya MouraAinda não há avaliações
- Uso da água ozonizada na medicinaDocumento29 páginasUso da água ozonizada na medicinaFabianaNisanAinda não há avaliações
- BioCel v2 03Documento11 páginasBioCel v2 03Vivi LeeAinda não há avaliações
- Hygirest ProprietariosDocumento170 páginasHygirest Proprietariosjo_ joaninha100% (3)
- Prova e Gabarito UEG 2018 2Documento19 páginasProva e Gabarito UEG 2018 2Pedro Henrique BorgesAinda não há avaliações
- Raiva Animal: 10 dias de observaçãoDocumento12 páginasRaiva Animal: 10 dias de observaçãoFelipeAinda não há avaliações
- BronchovaxomDocumento1 páginaBronchovaxomCharles FabioAinda não há avaliações
- Tesis Doctoral ConjuntivitisDocumento337 páginasTesis Doctoral ConjuntivitisPablo Del PozoAinda não há avaliações
- Aula 02 - Bioquímica Da CélulaDocumento27 páginasAula 02 - Bioquímica Da CélulaRenato ViniciusAinda não há avaliações
- Agrobactérias e BiotecnologiaDocumento52 páginasAgrobactérias e BiotecnologiaJoão DeganAinda não há avaliações
- Carbapenemases: Ameaça Global de ResistênciaDocumento33 páginasCarbapenemases: Ameaça Global de ResistênciaClaudio SoaresAinda não há avaliações
- Bolores conceito e característicasDocumento39 páginasBolores conceito e característicasErivan OliveiraAinda não há avaliações
- Questões de ImunologiaDocumento87 páginasQuestões de ImunologiaDudaBarbosaAinda não há avaliações
- Atlas de Microbiologia Compactado 2Documento32 páginasAtlas de Microbiologia Compactado 2Anine LeãoAinda não há avaliações