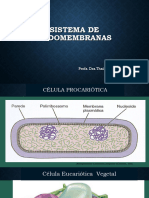
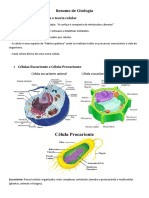
Você também pode gostar
- Tráfego Intracelular de VesículasDocumento5 páginasTráfego Intracelular de VesículasjbeloniAinda não há avaliações
- Resmo - Tráfego Intracelular de VesículasDocumento12 páginasResmo - Tráfego Intracelular de Vesículas9vvzhn8krvAinda não há avaliações
- Organelas e transporte de proteínasDocumento14 páginasOrganelas e transporte de proteínasRapha BuenoAinda não há avaliações
- Trafego Intracelular de VesiculasDocumento15 páginasTrafego Intracelular de VesiculasbatistapessoaiandraAinda não há avaliações
- Sintese e Secreção de MacromoléculasDocumento7 páginasSintese e Secreção de MacromoléculasPablo MonticeliAinda não há avaliações
- Aula 5 Tranporte VesicularDocumento36 páginasAula 5 Tranporte VesicularÉrica Boer GarciaAinda não há avaliações
- O transporte entre citosol e núcleoDocumento12 páginasO transporte entre citosol e núcleoMatheus Correa0% (1)
- Aula 5 - BiocelDocumento13 páginasAula 5 - Biocelliz de andradeAinda não há avaliações
- Aula 14-15-16 - Sistemas de Endomembranas e Transporte CelularDocumento65 páginasAula 14-15-16 - Sistemas de Endomembranas e Transporte CelularAlejandro Morales GomezAinda não há avaliações
- Resumo - Envoltório Nuclear 2Documento4 páginasResumo - Envoltório Nuclear 29vvzhn8krvAinda não há avaliações
- Sist - ENDOMEMBRANA 2023 - CBiomedDocumento46 páginasSist - ENDOMEMBRANA 2023 - CBiomedavenidadepirocaAinda não há avaliações
- Aula 4 - BiocelDocumento15 páginasAula 4 - Biocelliz de andradeAinda não há avaliações
- Membranas e transporte: tipos e mecanismosDocumento6 páginasMembranas e transporte: tipos e mecanismosSheila PinhoAinda não há avaliações
- Transporte de Proteínas para Organelas por Sinais de DistribuiçãoDocumento5 páginasTransporte de Proteínas para Organelas por Sinais de DistribuiçãoMacklecio OliveiraAinda não há avaliações
- Retculo Endoplasmtico e GolgiDocumento24 páginasRetculo Endoplasmtico e GolgiCamila MendesAinda não há avaliações
- Aula 4 Tranporte VesicularDocumento37 páginasAula 4 Tranporte VesicularAntonio Carlos Cruz Da SilvaAinda não há avaliações
- Aula 3 - RE e GolgiDocumento42 páginasAula 3 - RE e GolgiCamila HerculanoAinda não há avaliações
- Via secreção biologia celularDocumento35 páginasVia secreção biologia celularKarina Fernandes ValeAinda não há avaliações
- Atmr 09 BiologiaDocumento13 páginasAtmr 09 Biologiapd536121Ainda não há avaliações
- CélulaDocumento9 páginasCélulaMariana RochaAinda não há avaliações
- Tráfego VesicularDocumento15 páginasTráfego Vesicularanonimo anonimoAinda não há avaliações
- Complexo de Golgi e LisossomosDocumento105 páginasComplexo de Golgi e LisossomosBruna SerraAinda não há avaliações
- Estudo Dirigido - Reticulo Endoplasmático e LisossomosDocumento6 páginasEstudo Dirigido - Reticulo Endoplasmático e LisossomosRafael_F_AntunesAinda não há avaliações
- BIOLOGIA CELULAR MÓDULO 3 Video 1,2,3Documento29 páginasBIOLOGIA CELULAR MÓDULO 3 Video 1,2,3Maria lAinda não há avaliações
- Complexo de Golgi e LisossomosDocumento9 páginasComplexo de Golgi e LisossomosJúlia MottaAinda não há avaliações
- Membrana Celular: Proteínas e TransportesDocumento9 páginasMembrana Celular: Proteínas e TransportesAna Martins100% (1)
- Estrutura e função das membranas celulares e sistemas de transporteDocumento14 páginasEstrutura e função das membranas celulares e sistemas de transporteLuis RicardoAinda não há avaliações
- Revisão BIO CEL IDocumento27 páginasRevisão BIO CEL IgiovanaAinda não há avaliações
- Aula sobre endomembranas e síntese de proteínasDocumento2 páginasAula sobre endomembranas e síntese de proteínasAna Clara FrotaAinda não há avaliações
- Resumo da estrutura e função da célulaDocumento13 páginasResumo da estrutura e função da célulaLuciana Taliano50% (2)
- A Síntese Proteica No Retículo Endoplasmático RugosoDocumento1 páginaA Síntese Proteica No Retículo Endoplasmático RugosoHeloise PereiraAinda não há avaliações
- Estudo Dirigido - Compartimentos Intracelulares e Transporte de ProteínasDocumento2 páginasEstudo Dirigido - Compartimentos Intracelulares e Transporte de ProteínasLuiza LopesAinda não há avaliações
- As Sequências-Sinal Direcionam As Proteínas para Os Compartimentos CorretosDocumento4 páginasAs Sequências-Sinal Direcionam As Proteínas para Os Compartimentos CorretosRafael LuizAinda não há avaliações
- Obtenção de Matéria PDFDocumento15 páginasObtenção de Matéria PDFLeticia RussoAinda não há avaliações
- Apostila - MitocondriasDocumento3 páginasApostila - Mitocondriastamarauescbiomed931Ainda não há avaliações
- Compartimentos Intracelulares e TransporteDocumento8 páginasCompartimentos Intracelulares e Transportefevabax924Ainda não há avaliações
- Estudo Biologia2Documento9 páginasEstudo Biologia2WQWQWEQWEQAinda não há avaliações
- As Organelas Na Síntese e Degradação Das MoléculasDocumento8 páginasAs Organelas Na Síntese e Degradação Das MoléculasSamuel LemosAinda não há avaliações
- Aula 7 - GOLGI e Transporte VesicularDocumento26 páginasAula 7 - GOLGI e Transporte Vesicular18taisAinda não há avaliações
- Síntese proteica em 3 etapasDocumento11 páginasSíntese proteica em 3 etapasJoão SilvaAinda não há avaliações
- Fisiologia - ResumoDocumento5 páginasFisiologia - Resumolarissagabriela.limaAinda não há avaliações
- Biologia celular: Ciclo da vesícula sinápticaDocumento7 páginasBiologia celular: Ciclo da vesícula sinápticaJoyci StephanyAinda não há avaliações
- Aula 12 GolgiDocumento26 páginasAula 12 GolgiJulia Martins LotherAinda não há avaliações
- Funções organelas célula vegetal animalDocumento3 páginasFunções organelas célula vegetal animalThaís LuzAinda não há avaliações
- Aula 1 - Membrana PlasmáticaDocumento50 páginasAula 1 - Membrana PlasmáticaCamila HerculanoAinda não há avaliações
- Introdução ao metabolismo celularDocumento35 páginasIntrodução ao metabolismo celularvinicius dinizAinda não há avaliações
- Via Secretória e OrganelosDocumento12 páginasVia Secretória e Organelosjoaopinheiro88Ainda não há avaliações
- FISIOLOGIA MatériaDocumento151 páginasFISIOLOGIA MatériaSarah Livramento ZampirolliAinda não há avaliações
- CEDERJ Biologia Celular I - Gabarito 13 A 20Documento9 páginasCEDERJ Biologia Celular I - Gabarito 13 A 20api-3800070Ainda não há avaliações
- Bio Cel Aula 3Documento5 páginasBio Cel Aula 3JoséAinda não há avaliações
- Roteiro P3 - Semestre 2021Documento23 páginasRoteiro P3 - Semestre 2021Ediane RubasAinda não há avaliações
- Estudo Dirigido de Membranas A NeurotransmissoresDocumento4 páginasEstudo Dirigido de Membranas A Neurotransmissoreskarina garzAinda não há avaliações
- Resumo PsicofármacoDocumento7 páginasResumo PsicofármacoRianne GomesAinda não há avaliações
- Exercicio Citologia RespondidoDocumento10 páginasExercicio Citologia RespondidoWanessa TeófilaAinda não há avaliações
- Mitocôndria: estrutura, função e transporte de proteínasDocumento8 páginasMitocôndria: estrutura, função e transporte de proteínasMaíra SermidiAinda não há avaliações
- Resumo I (Continuação)Documento20 páginasResumo I (Continuação)Bento BentoAinda não há avaliações
- REL e RERDocumento12 páginasREL e RERAmanda FariaAinda não há avaliações
- 21 Pop Limpeza e Desinfecção Do Aparelho de Ultrassom e Esterilização e Armazenamento Dos AcessóriosDocumento2 páginas21 Pop Limpeza e Desinfecção Do Aparelho de Ultrassom e Esterilização e Armazenamento Dos Acessóriosvisiomed100% (1)
- Ficha de Informação sobre Álcool IsopropílicoDocumento3 páginasFicha de Informação sobre Álcool IsopropílicoAdilson VantineAinda não há avaliações
- Viscosidade de magmas e tipos de vulcõesDocumento3 páginasViscosidade de magmas e tipos de vulcõesPamela CarmoAinda não há avaliações
- QUI102 Ex 01 Lucas Paceli Cap1 2 3 4Documento26 páginasQUI102 Ex 01 Lucas Paceli Cap1 2 3 4Lucas PaceliAinda não há avaliações
- Cuidados com Preparações InjectáveisDocumento10 páginasCuidados com Preparações Injectáveisrogerio Jose SobraAinda não há avaliações
- Carbohidratos-definição,funções,classificação,metabolismoDocumento48 páginasCarbohidratos-definição,funções,classificação,metabolismoemile sousaAinda não há avaliações
- Fispq Pigmento Plastisol Verde BandeiraDocumento8 páginasFispq Pigmento Plastisol Verde BandeiraDouglas PedronAinda não há avaliações
- Aula Prática - Determinação de Cloretos em ÁguaDocumento3 páginasAula Prática - Determinação de Cloretos em ÁguaAntonioBertoldoAinda não há avaliações
- Mini Curso de Lubrificação NSKDocumento63 páginasMini Curso de Lubrificação NSKalex.mtsmarquesAinda não há avaliações
- Respiração Aeróbia 10º AnoDocumento19 páginasRespiração Aeróbia 10º AnoLourenço RaposoAinda não há avaliações
- Aminoácidos essenciais para síntese de proteínasDocumento2 páginasAminoácidos essenciais para síntese de proteínasAna Julia De Almeida BragaAinda não há avaliações
- Aula 1 - 15 de Fevereiro de 2021Documento84 páginasAula 1 - 15 de Fevereiro de 2021Carolina PintoAinda não há avaliações
- Baú Do Charcuteiro 220720Documento16 páginasBaú Do Charcuteiro 220720Diego RibeiroAinda não há avaliações
- Ácido 5 - Sulfosalicílico PaDocumento7 páginasÁcido 5 - Sulfosalicílico PaCarollina de Abrantes CasaAinda não há avaliações
- Inventário de Produtos Químicos ExemploDocumento2 páginasInventário de Produtos Químicos ExemploLídia Marina Torres CarvalhoAinda não há avaliações
- Modelo-Certificado Treinamento Sinalização e Manuseio Químicos - DIEGO OVERNECK MARQUES 07.10.2021Documento2 páginasModelo-Certificado Treinamento Sinalização e Manuseio Químicos - DIEGO OVERNECK MARQUES 07.10.2021VALÉRIA PEREIRA ViannaAinda não há avaliações
- Apresentação - MembranasDocumento178 páginasApresentação - MembranasElvis GuimaraesAinda não há avaliações
- Utilizao de Resduos Madeireiros e Plstico Ps-Consumo Como Alternativa para Produo de Wood Plastic CoDocumento5 páginasUtilizao de Resduos Madeireiros e Plstico Ps-Consumo Como Alternativa para Produo de Wood Plastic CoAndreaAinda não há avaliações
- Lista de Sobre Teoria Atómica e Estrutura Da Materia, Ligação Química e Termodinâmica QuímicaDocumento6 páginasLista de Sobre Teoria Atómica e Estrutura Da Materia, Ligação Química e Termodinâmica QuímicaRafael faruqueAinda não há avaliações
- Guia de Produtos Verso DigitalDocumento220 páginasGuia de Produtos Verso DigitalVanessa rosa dos santosAinda não há avaliações
- Estrutura e Função das CélulasDocumento32 páginasEstrutura e Função das CélulasGEOVANNA MARIA DE MATOSAinda não há avaliações
- Avaliação de Química Geral e OrgânicaDocumento4 páginasAvaliação de Química Geral e OrgânicaFernandaAinda não há avaliações
- Leitura Complementar - RessonânciaDocumento3 páginasLeitura Complementar - RessonânciaRayane TeodoroAinda não há avaliações
- Tratamentos térmicos: aplicações e processosDocumento73 páginasTratamentos térmicos: aplicações e processosRafael OliveiraAinda não há avaliações
- Relatório de CinzasDocumento6 páginasRelatório de CinzasPaula Keiko PeraltaAinda não há avaliações
- Aula 03 - Membrana Celular e Comunicação CelularDocumento148 páginasAula 03 - Membrana Celular e Comunicação CelularthamiAinda não há avaliações
- Óleo Essencial PDFDocumento6 páginasÓleo Essencial PDFJohann Costa CastelliAinda não há avaliações
- Checklist Capacetes - MSADocumento11 páginasChecklist Capacetes - MSADANIELAinda não há avaliações
- Deiton Traktor Transmission SAE 10W API MT-1 PDFDocumento10 páginasDeiton Traktor Transmission SAE 10W API MT-1 PDFAndre CruzAinda não há avaliações
- Exercicios Eletroquimica Coc ResolveDocumento2 páginasExercicios Eletroquimica Coc ResolveRaphael Rodrigues CostaAinda não há avaliações