Você também pode gostar
- O ciclo celular: divisão e renovaçãoDocumento3 páginasO ciclo celular: divisão e renovaçãoMarta OliveiraAinda não há avaliações
- 1.2 - Ciclo CelularDocumento77 páginas1.2 - Ciclo Celulartiagocostinha1904Ainda não há avaliações
- 03 componentes celularesDocumento6 páginas03 componentes celularesJandderson CostaAinda não há avaliações
- Núcleo Celular: FunçãoDocumento4 páginasNúcleo Celular: FunçãoCowardAinda não há avaliações
- Ciclo Celular - 2023 - 2024Documento30 páginasCiclo Celular - 2023 - 2024beatrizmhpereiraAinda não há avaliações
- 1.2 - Ciclo CelularDocumento75 páginas1.2 - Ciclo CelularCláudia PereiraAinda não há avaliações
- Resumo Genética (1º Período - 1º Ciclo)Documento9 páginasResumo Genética (1º Período - 1º Ciclo)laurafsp.brAinda não há avaliações
- Ciclo CelularDocumento48 páginasCiclo CelularMiguelAinda não há avaliações
- DNA, RNA e a replicação celularDocumento11 páginasDNA, RNA e a replicação celularGleyse GomesAinda não há avaliações
- Ciclo CelularDocumento4 páginasCiclo Celular6mqjp5hhsfAinda não há avaliações
- 5 - Ciclo CelularDocumento60 páginas5 - Ciclo Celularana carolAinda não há avaliações
- Ciclo celular e divisão celularDocumento38 páginasCiclo celular e divisão celularSofiaAinda não há avaliações
- Biologia - Ciclo CelularDocumento4 páginasBiologia - Ciclo CelularJoão AzevedoAinda não há avaliações
- Ciclo celular e divisão mitóticaDocumento19 páginasCiclo celular e divisão mitóticawesley harrisonAinda não há avaliações
- II - Ciclo CelularDocumento20 páginasII - Ciclo CelularMagda RodriguesAinda não há avaliações
- Bio Campbell-1-500-151-200Documento50 páginasBio Campbell-1-500-151-200AsukaAinda não há avaliações
- 200-380-1-181Documento181 páginas200-380-1-181Renata RamosAinda não há avaliações
- Organização e Regulação Do Material GenéticoDocumento23 páginasOrganização e Regulação Do Material GenéticoLuana OliveiraAinda não há avaliações
- Citologia e Histologia - Aula 03Documento26 páginasCitologia e Histologia - Aula 03MAYARA FIALHO LEITEAinda não há avaliações
- 15 Núcleo CelularDocumento5 páginas15 Núcleo CelularSamuelSamuelAinda não há avaliações
- Estrutura e Funções do Núcleo CelularDocumento6 páginasEstrutura e Funções do Núcleo CelularJosé FernandesAinda não há avaliações
- 5 - Organização e Regulação Do Material GenéticoDocumento23 páginas5 - Organização e Regulação Do Material GenéticomarianaAinda não há avaliações
- DIVISÃO CELULAR - MitoseDocumento17 páginasDIVISÃO CELULAR - MitoseRauan BrendonAinda não há avaliações
- Núcleo e Ciclo CelularDocumento39 páginasNúcleo e Ciclo CelularLizAinda não há avaliações
- Ciclo celular: divisão e replicação do DNADocumento2 páginasCiclo celular: divisão e replicação do DNAMaria Abrantes TavaresAinda não há avaliações
- CromossomasDocumento3 páginasCromossomasDavid FernandesAinda não há avaliações
- Ciclo Celular: Divisão e Crescimento das CélulasDocumento6 páginasCiclo Celular: Divisão e Crescimento das CélulasFo oda-seAinda não há avaliações
- Aula 10 - Ciclo Celular e MitoseDocumento48 páginasAula 10 - Ciclo Celular e MitoseLucio MotaAinda não há avaliações
- Alteração genéticaDocumento6 páginasAlteração genéticaCarolina JorgeAinda não há avaliações
- Aula 10 - Ciclo Celular e MitoseDocumento48 páginasAula 10 - Ciclo Celular e Mitoseisabelnhamue175Ainda não há avaliações
- Morfologia celularDocumento21 páginasMorfologia celularEllen GonçalvesAinda não há avaliações
- Ciclo CelularDocumento9 páginasCiclo CelularMatilde De Matos GomesAinda não há avaliações
- 2 BiologiaDocumento12 páginas2 BiologiaMaria GonçalvesAinda não há avaliações
- Constituição de Um Cromossoma EucarióticoDocumento4 páginasConstituição de Um Cromossoma EucarióticoFarley AlvesAinda não há avaliações
- Apresent Ciclo-Celular Diferenciacao-CelularDocumento18 páginasApresent Ciclo-Celular Diferenciacao-CelularPokitasAinda não há avaliações
- Guião Organização Material GenéticoDocumento2 páginasGuião Organização Material GenéticoOlá AdeusAinda não há avaliações
- Ciclo CelularDocumento43 páginasCiclo CelularFrancisca PatameiraAinda não há avaliações
- Quanndo e Como Surgiram As CelulasDocumento3 páginasQuanndo e Como Surgiram As CelulasJoel Júnior SilvaAinda não há avaliações
- Aula 2 - Núcleo Celular 2021.2Documento32 páginasAula 2 - Núcleo Celular 2021.2Karvs100% (1)
- Apostila de Núcleo e Divisões CelularesDocumento22 páginasApostila de Núcleo e Divisões CelularesIara MardoiAinda não há avaliações
- Mapa MentalDocumento1 páginaMapa MentalevelyndasilvaaceniAinda não há avaliações
- Núcleo Celular e CromossomosDocumento4 páginasNúcleo Celular e CromossomosSuarezinho100% (8)
- Mitose Meiose - RemovedDocumento24 páginasMitose Meiose - Removedalexfreitaslima375Ainda não há avaliações
- Ciclo Celular e Crossomas e MutacoesDocumento7 páginasCiclo Celular e Crossomas e Mutacoesbeatriz.almeida.ferreira2006Ainda não há avaliações
- s13 Ciclo CelularDocumento5 páginass13 Ciclo CelularJuan Diego Huaman RamirezAinda não há avaliações
- Mitose e Meiose e Sintese de ProteinasDocumento4 páginasMitose e Meiose e Sintese de Proteinas457742Ainda não há avaliações
- Silo - Tips As Bases Citologicas Da HereditariedadeDocumento32 páginasSilo - Tips As Bases Citologicas Da HereditariedadeLuciano Pereira NascimentoAinda não há avaliações
- Resumo CitologiaDocumento10 páginasResumo CitologiaMaria VitoriaAinda não há avaliações
- Divisão Celular e CrescimentoDocumento40 páginasDivisão Celular e CrescimentotrilibiteAinda não há avaliações
- Núcleo Celular: Brenda de Oliveira Cristina GarciaDocumento40 páginasNúcleo Celular: Brenda de Oliveira Cristina GarciaGiovanna JoiceAinda não há avaliações
- Biologia da célulaDocumento52 páginasBiologia da célulaFrancisca Layslane Taveira dos AnjosAinda não há avaliações
- Aula 12 MitosemeioseDocumento11 páginasAula 12 MitosemeioseTalles Renato dos Santos PereiraAinda não há avaliações
- Introdução à Citogenética HumanaDocumento16 páginasIntrodução à Citogenética HumanaEVERTONAinda não há avaliações
- Estudo Dirigido Prova BiocelDocumento5 páginasEstudo Dirigido Prova BiocelJoão Vitor AlvesAinda não há avaliações
- NÚCLEODocumento7 páginasNÚCLEOAlineAinda não há avaliações
- Cromatina, Cromossomos e Divisão CelularDocumento7 páginasCromatina, Cromossomos e Divisão CelularMatheus GuimaraesAinda não há avaliações
- BIOLOGIADocumento15 páginasBIOLOGIAArthur SantosAinda não há avaliações
- Estrutura Cromossômica CelularDocumento1 páginaEstrutura Cromossômica CelularVera MenezesAinda não há avaliações
- EX Port639 F1 2021 V1 - NetDocumento8 páginasEX Port639 F1 2021 V1 - NetAntónio LucasAinda não há avaliações
- 17 07 2023 Comprovativo MatriculaDocumento5 páginas17 07 2023 Comprovativo MatriculaAntónio LucasAinda não há avaliações
- Importância Biológica da Mitose na Multiplicação e Regeneração CelularDocumento53 páginasImportância Biológica da Mitose na Multiplicação e Regeneração CelularAntónio LucasAinda não há avaliações
- AL1-Ciclo Do CobreDocumento13 páginasAL1-Ciclo Do CobreAntónio LucasAinda não há avaliações
- AlgoritmiaDocumento19 páginasAlgoritmiaAntónio LucasAinda não há avaliações
- A Queda de um Anjo por Camilo Castelo BrancoDocumento5 páginasA Queda de um Anjo por Camilo Castelo BrancoAntónio LucasAinda não há avaliações
- 6 A - Reprodução Nos Seres Vivos - Reprodução AssexuadaDocumento33 páginas6 A - Reprodução Nos Seres Vivos - Reprodução AssexuadaAntónio LucasAinda não há avaliações
- Melhoria das vias e segurança no Pinhal InteriorDocumento2 páginasMelhoria das vias e segurança no Pinhal InteriorAntónio LucasAinda não há avaliações
- Guia 5Documento1 páginaGuia 5António LucasAinda não há avaliações
- Guia 1Documento1 páginaGuia 1António LucasAinda não há avaliações
- CDS Propostas (NP)Documento7 páginasCDS Propostas (NP)António LucasAinda não há avaliações
- Os desafios do Distrito de Leiria: custos energéticos, fiscalidade e saúdeDocumento14 páginasOs desafios do Distrito de Leiria: custos energéticos, fiscalidade e saúdeAntónio LucasAinda não há avaliações
- Teste Sermc3a3o Sto Ant Aos Peixes PerguntasDocumento8 páginasTeste Sermc3a3o Sto Ant Aos Peixes PerguntasMaxim MarcuAinda não há avaliações
- Análise de Jogadores para o Meio-Campo do BenficaDocumento441 páginasAnálise de Jogadores para o Meio-Campo do BenficaAntónio LucasAinda não há avaliações
- Observação microscópica de células em meioseDocumento10 páginasObservação microscópica de células em meioseAntónio LucasAinda não há avaliações
- Pagamento Execução FiscalDocumento1 páginaPagamento Execução FiscalAntónio LucasAinda não há avaliações
- Células vegetais em ciclo celularDocumento10 páginasCélulas vegetais em ciclo celularAntónio LucasAinda não há avaliações
- Portugueses e a baixa produtividade: causas e soluçõesDocumento7 páginasPortugueses e a baixa produtividade: causas e soluçõesAntónio LucasAinda não há avaliações
- Nova PlaylistDocumento1 páginaNova PlaylistAntónio LucasAinda não há avaliações
- Pagamento Execução FiscalDocumento1 páginaPagamento Execução FiscalAntónio LucasAinda não há avaliações
- Estudo da constituição de uma florDocumento9 páginasEstudo da constituição de uma florAntónio LucasAinda não há avaliações
- Benefícios vulcões agricultura solo acne energia geotérmicaDocumento3 páginasBenefícios vulcões agricultura solo acne energia geotérmicaAntónio LucasAinda não há avaliações
- Escola Secundária Pedro NunesDocumento7 páginasEscola Secundária Pedro NunesAntónio LucasAinda não há avaliações
- Biodiversidade e célula: unidade estrutural e funcionalDocumento15 páginasBiodiversidade e célula: unidade estrutural e funcionalAntónio LucasAinda não há avaliações
- 2A. - Composição Química e Estrutura Dos Ácidos NucleicosDocumento15 páginas2A. - Composição Química e Estrutura Dos Ácidos NucleicosAntónio LucasAinda não há avaliações
- 3 - Sintese ProteicaDocumento34 páginas3 - Sintese ProteicaAntónio LucasAinda não há avaliações
- Escola Secundária Pedro Nune1Documento10 páginasEscola Secundária Pedro Nune1António LucasAinda não há avaliações
- 2 - Composição Química e Estrutura Dos Ácidos NucleicosDocumento20 páginas2 - Composição Química e Estrutura Dos Ácidos NucleicosAntónio LucasAinda não há avaliações
- Formação de Um Nucleótido e de Cadeia PolinucleotídicaDocumento3 páginasFormação de Um Nucleótido e de Cadeia PolinucleotídicaAntónio LucasAinda não há avaliações
- Anatomia (Introdução)Documento1 páginaAnatomia (Introdução)Hellen Santos BarbosaAinda não há avaliações
- Influência da massagem modeladora no tratamento do fibro edema gelóideDocumento17 páginasInfluência da massagem modeladora no tratamento do fibro edema gelóideAndréa F. S. LocarnoAinda não há avaliações
- Esqueleto Axial - Toda MatériaDocumento5 páginasEsqueleto Axial - Toda MatériaThomäs ShälëAinda não há avaliações
- ALDA CapsulaDocumento8 páginasALDA CapsulaJose FariaAinda não há avaliações
- Atlas Básico de HistologiaDocumento14 páginasAtlas Básico de HistologiaRaphaelEPCArAinda não há avaliações
- Sistema endócrino e seus principais hormôniosDocumento35 páginasSistema endócrino e seus principais hormôniosBARBARA MACHADO GARCIA MEDICINAAinda não há avaliações
- PAAF/BAAF para diagnóstico de lesões mamárias e tireoideiasDocumento12 páginasPAAF/BAAF para diagnóstico de lesões mamárias e tireoideiasfatyene FariasAinda não há avaliações
- Endocrino HipofiseDocumento59 páginasEndocrino HipofisewesleyfisioAinda não há avaliações
- Anatomia Topográfica Do Casco EquinoDocumento6 páginasAnatomia Topográfica Do Casco EquinoAlexandro SilvaAinda não há avaliações
- Sistema CirculatórioDocumento3 páginasSistema CirculatórioFernanda DamasioAinda não há avaliações
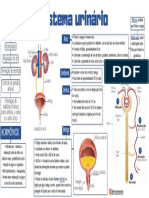
- Dois rins e seus componentes do sistema urinárioDocumento1 páginaDois rins e seus componentes do sistema urinárioJulio SanchezAinda não há avaliações
- Anatomia comparada Cão Equino RuminanteDocumento8 páginasAnatomia comparada Cão Equino Ruminantebhanny2Ainda não há avaliações
- Desenvolvimento do sistema digestórioDocumento23 páginasDesenvolvimento do sistema digestórioTin Tin GuerraAinda não há avaliações
- Doenças Humanas Associadas À Forma e Função Do Complexo de Golgi TraduzidoDocumento6 páginasDoenças Humanas Associadas À Forma e Função Do Complexo de Golgi TraduzidoMatheus RicardoAinda não há avaliações
- P1 - BiocelDocumento15 páginasP1 - BiocelEdiane RubasAinda não há avaliações
- Relatório cirurgia pescoço menina 5 anosDocumento1 páginaRelatório cirurgia pescoço menina 5 anosIvy CollyerAinda não há avaliações
- Reprodução humana e manipulação da fertilidadeDocumento8 páginasReprodução humana e manipulação da fertilidadeMariana RochaAinda não há avaliações
- Fisiologia Cardíaca: Sistema CirculatórioDocumento13 páginasFisiologia Cardíaca: Sistema CirculatórioEduardo RAinda não há avaliações
- Reprodução Humana PDFDocumento234 páginasReprodução Humana PDFPlinio ReisAinda não há avaliações
- Célula VegetalDocumento21 páginasCélula VegetalAdrielle JordanaAinda não há avaliações
- Cópia de Rinoplastia - Série Atlas de Cirurgia Plástica - TAUB, Peter J. BAKER, Stephen B. OCRDocumento163 páginasCópia de Rinoplastia - Série Atlas de Cirurgia Plástica - TAUB, Peter J. BAKER, Stephen B. OCRvaleriacampagnolo97Ainda não há avaliações
- Anatomia e fisiologia do sistema digestivo bovinoDocumento6 páginasAnatomia e fisiologia do sistema digestivo bovinoNorian Luis De Castro SiliprandiAinda não há avaliações
- Biologia SecundáriaDocumento27 páginasBiologia SecundáriaAlex MonitoAinda não há avaliações
- Embriologia TrabalhoDocumento6 páginasEmbriologia TrabalhoDedaldino Guer Panda GuerraAinda não há avaliações
- Neuroanatomia e vascularização cerebralDocumento21 páginasNeuroanatomia e vascularização cerebralNathalia WesslerAinda não há avaliações
- Sistema Digestório: Processos e EstruturaDocumento37 páginasSistema Digestório: Processos e EstruturaighormtAinda não há avaliações
- Anatomia Humana EssencialDocumento33 páginasAnatomia Humana EssencialEloiza AlmeidaAinda não há avaliações
- Sistema circulatório - IIDocumento17 páginasSistema circulatório - IIMaria Eduarda Araújo AquinoAinda não há avaliações
- Segunda Semana Do Desenvolvimento Embrionário Aula 04Documento12 páginasSegunda Semana Do Desenvolvimento Embrionário Aula 04Fabíula OrtegaAinda não há avaliações
- Aula 2 - Anatomia e Sinais VitaisDocumento64 páginasAula 2 - Anatomia e Sinais Vitaisemilly freitasAinda não há avaliações