Você também pode gostar
- DorDocumento58 páginasDorDesireGarcia100% (5)
- Neuroanatomia Da DorDocumento5 páginasNeuroanatomia Da DorRoberto HangleyAinda não há avaliações
- Sistema Nervoso-ApresentaçãoDocumento3 páginasSistema Nervoso-Apresentaçãotatiana.barroqueirogmail.com barroqueiroAinda não há avaliações
- Dor ResumoDocumento4 páginasDor ResumoGee AndlerAinda não há avaliações
- DORDocumento5 páginasDORSoraia GonçalvesAinda não há avaliações
- A Fisiologia Da Dor: IntroduçãoDocumento12 páginasA Fisiologia Da Dor: Introduçãoesmael franciscoAinda não há avaliações
- Dor, Cefaleia e Sensações TérmicasDocumento27 páginasDor, Cefaleia e Sensações TérmicasBruno Fernandes BorginhoAinda não há avaliações
- Aula 2 Parte I - Nocicepção e TemperaturaDocumento15 páginasAula 2 Parte I - Nocicepção e TemperaturaPatrícia SilvaAinda não há avaliações
- Dor - Anatomofisiologia e MecanismosDocumento51 páginasDor - Anatomofisiologia e MecanismosWallberthy Linhares100% (1)
- Fisiopatologia Da Sensibilidade e Da DorDocumento47 páginasFisiopatologia Da Sensibilidade e Da Dorpereira_wilker9835100% (1)
- Neurofisiologia Aula 3Documento60 páginasNeurofisiologia Aula 3Maria Clara Figueira BarbosaAinda não há avaliações
- Nociceptores e Hiperalgesia.Documento32 páginasNociceptores e Hiperalgesia.Kedma VitóriaAinda não há avaliações
- Visao Geral Sistema SensitivoDocumento38 páginasVisao Geral Sistema SensitivoPriscilla Santos100% (1)
- Controle Da DorDocumento72 páginasControle Da DorSerJauM LoBauMAinda não há avaliações
- Capítulo 49 - Sensações Somáticas - Ian BezerraDocumento1 páginaCapítulo 49 - Sensações Somáticas - Ian Bezerrarbatalha2000Ainda não há avaliações
- Capítulo 01 - Aspectos Anatômicos e FisiopatológicosDocumento7 páginasCapítulo 01 - Aspectos Anatômicos e FisiopatológicosSandro PerilloAinda não há avaliações
- Resumo de DorDocumento11 páginasResumo de DorLucianaAinda não há avaliações
- Neurônios, Sinapses e DorDocumento25 páginasNeurônios, Sinapses e DorErika BarretoAinda não há avaliações
- P1 TutoriaDocumento6 páginasP1 TutoriaMaria Eduarda GeneraliAinda não há avaliações
- Fisiologia Da DorDocumento35 páginasFisiologia Da DorDEBORA CAPODALIOAinda não há avaliações
- Neurofisiologia Sensitiva e MotoraDocumento8 páginasNeurofisiologia Sensitiva e MotoraJaqueline Dias100% (1)
- Sensacoes ExteroceptivasDocumento66 páginasSensacoes Exteroceptivasjorgelina_riveraAinda não há avaliações
- Dor e TemperaturaDocumento3 páginasDor e TemperaturaANNA BEATRIZ MINGA ZANARDIAinda não há avaliações
- NocicepçãoDocumento17 páginasNocicepçãoaron staffenAinda não há avaliações
- Neurofisiologia Da DorDocumento7 páginasNeurofisiologia Da DorJoao MathiasAinda não há avaliações
- Capítulo 48 - Sensações Somáticas Ii - Dor, Cefaléia e Sensações Térmicas - 3 PáginasDocumento3 páginasCapítulo 48 - Sensações Somáticas Ii - Dor, Cefaléia e Sensações Térmicas - 3 Páginasandreamokada100% (1)
- Fisioterapia o Controle Da DorDocumento4 páginasFisioterapia o Controle Da DorFran2502Ainda não há avaliações
- Captura de Tela 2020-11-18 À(s) 11.41.33Documento47 páginasCaptura de Tela 2020-11-18 À(s) 11.41.33Angelo TorresAinda não há avaliações
- Sensações SomáticasDocumento4 páginasSensações SomáticasPAULO VINICIUS DOS SANTOS CHAGASAinda não há avaliações
- Diferença Na Estrutura (Fibra Adelta E C)Documento5 páginasDiferença Na Estrutura (Fibra Adelta E C)Giovanna Hessmann FernandesAinda não há avaliações
- Resumo Vias Aferentes Medulares - Dor e TemperaturaDocumento3 páginasResumo Vias Aferentes Medulares - Dor e TemperaturaIsabella Nino100% (1)
- Resumão Tuts Uc11Documento52 páginasResumão Tuts Uc11Fernanda Eder FerreiraAinda não há avaliações
- Resumo de Conceitos Básicos em EletrotermofototerapiaDocumento13 páginasResumo de Conceitos Básicos em EletrotermofototerapiaCanal MagnoBaits PescariasAinda não há avaliações
- Vias AscendentesDocumento9 páginasVias AscendentesLucianaAinda não há avaliações
- Dor e AnalgesiaDocumento7 páginasDor e AnalgesiaLara TavaresAinda não há avaliações
- 6 Texto - Os Sentidos SomáticosDocumento4 páginas6 Texto - Os Sentidos SomáticosCoordenação BiomedicinaAinda não há avaliações
- Aula 01. Dor Pedro Paul 07.02Documento13 páginasAula 01. Dor Pedro Paul 07.02Diana Myrria OliveiraAinda não há avaliações
- Aula - Dor - Bases NeuraisDocumento13 páginasAula - Dor - Bases NeuraissimedtocantinsAinda não há avaliações
- FisiologiaDocumento2 páginasFisiologiaIzabele AmandaAinda não há avaliações
- Relatório Fisiologia Sistema SensorialDocumento17 páginasRelatório Fisiologia Sistema SensorialArquimeniaAinda não há avaliações
- Taxonomia Da DorDocumento38 páginasTaxonomia Da DorrafadxpqAinda não há avaliações
- ApostilaDocumento14 páginasApostilaAnderson LimaAinda não há avaliações
- Trilha Semiologia Da DorDocumento12 páginasTrilha Semiologia Da DorCAMILA EUQUERES PARTATAAinda não há avaliações
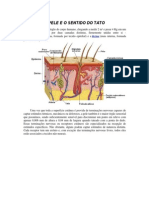
- A Pele e o Sentido Do TatoDocumento5 páginasA Pele e o Sentido Do TatotatistapAinda não há avaliações
- Abordagem SensorialDocumento7 páginasAbordagem SensorialReinaldo Dos AnjosAinda não há avaliações
- Agulhamento A SecoDocumento41 páginasAgulhamento A SecoYara Borini RossiAinda não há avaliações
- Sensações Somáticas II - DorDocumento4 páginasSensações Somáticas II - DorMariana PerezAinda não há avaliações
- NocicepçãoDocumento3 páginasNocicepçãoPedro NunoAinda não há avaliações
- Fisiopatologia Da: @ka - EnfermeiraDocumento16 páginasFisiopatologia Da: @ka - EnfermeiraBianca Mara CristanteAinda não há avaliações
- Dor AbdominalDocumento7 páginasDor AbdominalGabriel CagninAinda não há avaliações
- Fisiologia Da Dor PDFDocumento3 páginasFisiologia Da Dor PDFGrisel Ibáñez100% (1)
- HarrisonDocumento1.351 páginasHarrisonMaria LimaAinda não há avaliações
- Harisson DorDocumento113 páginasHarisson DorRangellwlAinda não há avaliações
- Usp - Propriocepção, Nocicepcão, ExterocepçãoDocumento33 páginasUsp - Propriocepção, Nocicepcão, ExterocepçãoLaura Schulz100% (1)
- Questões Fisio Miv 8Documento7 páginasQuestões Fisio Miv 8Amanda AAinda não há avaliações
- Semiologia Da DorDocumento12 páginasSemiologia Da DorJoão PauloAinda não há avaliações
- Avaliação do Controle Autonômico Baseada na Análise da Variabilidade da Frequência Cardíaca: Proposta de Método Otimizado para Ativação Vagal e Aplicação da Ferramenta para Estimativa da Dor em Intervenções ClínicasNo EverandAvaliação do Controle Autonômico Baseada na Análise da Variabilidade da Frequência Cardíaca: Proposta de Método Otimizado para Ativação Vagal e Aplicação da Ferramenta para Estimativa da Dor em Intervenções ClínicasAinda não há avaliações
- Temática - Ciências da Vida - Relação e CoordenaçãoNo EverandTemática - Ciências da Vida - Relação e CoordenaçãoAinda não há avaliações
- Dor 4Documento3 páginasDor 4Anuah Mansur AbudAinda não há avaliações
- Dor 3Documento11 páginasDor 3Anuah Mansur AbudAinda não há avaliações
- Módulo I - Dor Problema 2 - Assombração Objetivos: Lorena de Souza Santos Medicina - Turma XX 5 Etapa - 2º Semestre 2015Documento6 páginasMódulo I - Dor Problema 2 - Assombração Objetivos: Lorena de Souza Santos Medicina - Turma XX 5 Etapa - 2º Semestre 2015Anuah Mansur AbudAinda não há avaliações
- ANAMNESE GinecologicaDocumento3 páginasANAMNESE GinecologicaAnuah Mansur AbudAinda não há avaliações
- Caderno de Questões - FilosofiaDocumento5 páginasCaderno de Questões - FilosofiaRogério GonçalvesAinda não há avaliações
- Lista 1Documento6 páginasLista 1Ricardo de AlmeidaAinda não há avaliações
- Jacques MaritainDocumento3 páginasJacques MaritainGladius100% (1)
- Fagifor. FortalezaDocumento61 páginasFagifor. FortalezaMécia costaAinda não há avaliações
- Modelo de Currículo para Vaga de Estágio em DireitoDocumento1 páginaModelo de Currículo para Vaga de Estágio em DireitoRamon SouzaAinda não há avaliações
- Folder MedilabDocumento12 páginasFolder MedilabHerick SavioneAinda não há avaliações
- BT-VRV-022 (Anexo) Daikin + AAF 20220727Documento10 páginasBT-VRV-022 (Anexo) Daikin + AAF 20220727RODOLFO ARTHURAinda não há avaliações
- Estudo para Cálculo Do Número de Curva Do Método SCS para A Região Norte Da Bacia Hidrográfica Do Rio CachoeiraDocumento57 páginasEstudo para Cálculo Do Número de Curva Do Método SCS para A Região Norte Da Bacia Hidrográfica Do Rio CachoeiraMariana GonçalvesAinda não há avaliações
- Gerenciamento de ProjetosDocumento94 páginasGerenciamento de ProjetosGustavoAinda não há avaliações
- Especificações Técnicas para Quadro de Comando de Motores - QCMDocumento4 páginasEspecificações Técnicas para Quadro de Comando de Motores - QCMotavianoAinda não há avaliações
- Phill Kotler: - Marketing Lateral E VerticalDocumento35 páginasPhill Kotler: - Marketing Lateral E VerticalMarcos ÁlvaresAinda não há avaliações
- Pi PR 034.2Documento1 páginaPi PR 034.2CHARLES FONSECAAinda não há avaliações
- Gabarito 05 - Exercicio de Fixação 05 - 2 Lei de Ohm Potencia e Enregia PDFDocumento8 páginasGabarito 05 - Exercicio de Fixação 05 - 2 Lei de Ohm Potencia e Enregia PDFDaniel Silva De OliveiraAinda não há avaliações
- Biometria Aplicada Ao Estudo Da Diversidade GenéticaDocumento626 páginasBiometria Aplicada Ao Estudo Da Diversidade Genética28rhq6mbfkAinda não há avaliações
- Relatorio 8 Final EletronicaDocumento3 páginasRelatorio 8 Final EletronicaPedro JacquesAinda não há avaliações
- Bicicleta Estrada Triban 500Documento4 páginasBicicleta Estrada Triban 500Antonio Mendes0% (1)
- TCC - Linux - Rodrigo Dall AlbaDocumento61 páginasTCC - Linux - Rodrigo Dall Albardallalba100% (1)
- Duque Da Meia-Noite - Tessa DareDocumento385 páginasDuque Da Meia-Noite - Tessa DareAna Ivone Salomon Marques100% (3)
- Realismo Naturalismo e ParnasianismoDocumento5 páginasRealismo Naturalismo e ParnasianismoLucieneAndreAinda não há avaliações
- Relatório de Estágio Obrigatório - Marcus v. de B. SilvaDocumento59 páginasRelatório de Estágio Obrigatório - Marcus v. de B. SilvaVinícius de BritoAinda não há avaliações
- Apostila - Varias Maneiras de Fotometrar PDFDocumento77 páginasApostila - Varias Maneiras de Fotometrar PDFJuliana Rangel BicudoAinda não há avaliações
- Catalogo CorfioDocumento28 páginasCatalogo CorfioJoão MenegazAinda não há avaliações
- RelatorioSituacaoFiscal 28755666000179 20230303Documento2 páginasRelatorioSituacaoFiscal 28755666000179 20230303Rogerio Donato PepeAinda não há avaliações
- Apostila de Administração Enfase em Departamento PessoalDocumento50 páginasApostila de Administração Enfase em Departamento Pessoalgabriela_saldan2552Ainda não há avaliações
- Habilidades para Uma Carreira de Sucesso Na EngenhariaDocumento5 páginasHabilidades para Uma Carreira de Sucesso Na EngenhariaAna Julia BicalhoAinda não há avaliações
- Hoops OnDocumento95 páginasHoops Onevellenaquino50Ainda não há avaliações
- Como Produzir Conservas Que Não Estragam - Antonino PaixãoDocumento64 páginasComo Produzir Conservas Que Não Estragam - Antonino PaixãoJosemar SilvaAinda não há avaliações
- Módulo 1 - Aspectos Gerais Sobre Os Indicadores de Desempenho InstitucionalDocumento10 páginasMódulo 1 - Aspectos Gerais Sobre Os Indicadores de Desempenho InstitucionalGean CarlosAinda não há avaliações
- Apostila Saúde OcupacionalDocumento200 páginasApostila Saúde OcupacionalBruno MelloAinda não há avaliações
- Modelo de Relatório Do Estágio CurricularDocumento11 páginasModelo de Relatório Do Estágio CurricularVictória SousaAinda não há avaliações