Você também pode gostar
- Síntese Aula 7 (Lipídios)Documento3 páginasSíntese Aula 7 (Lipídios)Julyana RochaAinda não há avaliações
- O Livro De Receitas Para Tudo Com Baixo Colesterol Id 3389063 Produtor RascunhoNo EverandO Livro De Receitas Para Tudo Com Baixo Colesterol Id 3389063 Produtor RascunhoAinda não há avaliações
- Metabolismo dos lipídeosDocumento23 páginasMetabolismo dos lipídeosYam VictorAinda não há avaliações
- Atenção Nutricional Na Cirurgia Bariátrica e MetabólicaNo EverandAtenção Nutricional Na Cirurgia Bariátrica e MetabólicaNota: 5 de 5 estrelas5/5 (1)
- Dislipidemias: conceito, epidemiologia e metabolismo lipídicoDocumento28 páginasDislipidemias: conceito, epidemiologia e metabolismo lipídicoRoberta Violante100% (1)
- Sistema de Pontos para Controle de Colesterol e Gordura no Sangue: Ajude o seu coração a viver mais e melhorNo EverandSistema de Pontos para Controle de Colesterol e Gordura no Sangue: Ajude o seu coração a viver mais e melhorAinda não há avaliações
- Bases Fisiopatologicas Da Dislipidemia AterogenicaDocumento7 páginasBases Fisiopatologicas Da Dislipidemia AterogenicaPedroAinda não há avaliações
- Relatorio de Dosagem de Triglicerides e As DislipidemiasDocumento11 páginasRelatorio de Dosagem de Triglicerides e As DislipidemiasJohnny Percussionista100% (3)
- Curso Técnico em Alimentos: Gorduras TransDocumento8 páginasCurso Técnico em Alimentos: Gorduras TransAna Paula GalvãoAinda não há avaliações
- Interpretação de exames de perfil lipídico e distúrbios metabólicosDocumento131 páginasInterpretação de exames de perfil lipídico e distúrbios metabólicosJefferson Peres de MacedoAinda não há avaliações
- Trabalho: DISLIPIDEMIASDocumento8 páginasTrabalho: DISLIPIDEMIASrafa.arruda2Ainda não há avaliações
- Bioquímica Clínica - Perfil Lipídico 2021Documento44 páginasBioquímica Clínica - Perfil Lipídico 2021Karina Felinto0% (1)
- Doseamento de LípidosDocumento4 páginasDoseamento de LípidosAlexandra MarquesAinda não há avaliações
- Ebook Gorduras e Saúde CardiovascularDocumento20 páginasEbook Gorduras e Saúde CardiovascularClézia CarvalhoAinda não há avaliações
- Estrutura e função dos principais lipídiosDocumento3 páginasEstrutura e função dos principais lipídiosPapelaria TeixeiraAinda não há avaliações
- E-Book Lipídios - NAPEDocumento26 páginasE-Book Lipídios - NAPEDiego SantosAinda não há avaliações
- LipídeosDocumento6 páginasLipídeosRogério RosaAinda não há avaliações
- Aula LipídeosDocumento44 páginasAula LipídeosVanessa Ferreira CostaAinda não há avaliações
- NUTRIÇÃO HUMANA I - Estudo Dirigido 2Documento3 páginasNUTRIÇÃO HUMANA I - Estudo Dirigido 2rafinha_mendesAinda não há avaliações
- Cap 4Documento29 páginasCap 4Juliana OliveiraAinda não há avaliações
- Lipídios 2021Documento51 páginasLipídios 2021Letícia Gabrielly PortoAinda não há avaliações
- Farmacologia M4. Parte II-1Documento22 páginasFarmacologia M4. Parte II-1JamanaAinda não há avaliações
- Dislipidemias e Prevenção Da AteroscleroseDocumento9 páginasDislipidemias e Prevenção Da AteroscleroseAlice MirandaAinda não há avaliações
- Cartilha DislipidemiaDocumento12 páginasCartilha DislipidemiaRayane Silva100% (1)
- Estudo Dirigido LipideosDocumento8 páginasEstudo Dirigido LipideosFernanda Aline da SilvaAinda não há avaliações
- Aula 2Documento36 páginasAula 2LuisAinda não há avaliações
- Metabolismo lipídico: uma visão geralDocumento37 páginasMetabolismo lipídico: uma visão geralMônica Amaral LenziAinda não há avaliações
- Relatório Oteosclerose LuanaDocumento7 páginasRelatório Oteosclerose LuanaLuanaAinda não há avaliações
- Questões ColesterolDocumento2 páginasQuestões ColesterolAndréia LaraAinda não há avaliações
- O mito das gorduras saturadasDocumento19 páginasO mito das gorduras saturadasTatiani SilvaAinda não há avaliações
- Dislipidemias e tratamentosDocumento11 páginasDislipidemias e tratamentosDanuzio SilvaAinda não há avaliações
- GordurasDocumento10 páginasGordurasCAPAINA BENCILARIO ANDRE FAinda não há avaliações
- Doenças cardiovasculares, metabolismo lipídeos e dislipidemiasDocumento2 páginasDoenças cardiovasculares, metabolismo lipídeos e dislipidemiasNatalia DiasAinda não há avaliações
- Lipídios e DislipidemiasDocumento40 páginasLipídios e DislipidemiasRonara GomesAinda não há avaliações
- Nutrição - I - Efeitos Das Gorduras No Organismo - I - Saúde em MovimentoDocumento4 páginasNutrição - I - Efeitos Das Gorduras No Organismo - I - Saúde em MovimentoSerraAinda não há avaliações
- Monografia AdeSDocumento20 páginasMonografia AdeSVanessa Gama GalassoAinda não há avaliações
- Prof. Arnaldo (1) Atualizado 28-11Documento11 páginasProf. Arnaldo (1) Atualizado 28-11antonio.lisboa20018Ainda não há avaliações
- Ômega-3: Benefícios e Fontes AlimentaresDocumento38 páginasÔmega-3: Benefícios e Fontes AlimentarescarlosAinda não há avaliações
- Bioquímica Aplicada À Saúde-Unidade 3-3.1-1ºsem2022Documento29 páginasBioquímica Aplicada À Saúde-Unidade 3-3.1-1ºsem2022Kauanna MartinsAinda não há avaliações
- DislipidemiasDocumento11 páginasDislipidemiasEvelyn385Ainda não há avaliações
- O Papel Dos Ácidos Graxos Na Prevenção E No Tratamento de Doenças CardiovascularesDocumento24 páginasO Papel Dos Ácidos Graxos Na Prevenção E No Tratamento de Doenças CardiovascularesJessica MonteiroAinda não há avaliações
- GlicerideosDocumento13 páginasGlicerideosAlfredo Zacarias Feduca AzcAinda não há avaliações
- Lipídios AulaDocumento38 páginasLipídios AulaBernadete AraújoAinda não há avaliações
- Pravastatina SodicaDocumento2 páginasPravastatina SodicacelmorcelliAinda não há avaliações
- Guia Nutricao em AIDS 02Documento40 páginasGuia Nutricao em AIDS 02Paula SandesAinda não há avaliações
- DislipidemiaDocumento31 páginasDislipidemiaJueson OlimpioAinda não há avaliações
- Lipídios: gorduras, óleos, ácidos graxos e funções biológicasDocumento14 páginasLipídios: gorduras, óleos, ácidos graxos e funções biológicasbrunaAinda não há avaliações
- O que é e faz o colesterolDocumento21 páginasO que é e faz o colesterolPaulo GomesAinda não há avaliações
- Aula 6_26_09_DislipidemiaDocumento67 páginasAula 6_26_09_DislipidemiaYam VictorAinda não há avaliações
- 3 Bioquímica Dos Lipídeos, Carboidratos, Vitaminas e MineraisDocumento44 páginas3 Bioquímica Dos Lipídeos, Carboidratos, Vitaminas e MineraisrilixgsimululadorAinda não há avaliações
- 26 1008 Gorduras TransDocumento12 páginas26 1008 Gorduras TransMARIA EDUARDA VIEIRA HOLTZAinda não há avaliações
- DislipidemiaDocumento17 páginasDislipidemiaAbel Chacanessa de JesusAinda não há avaliações
- Dislipidemia: o que é, causas e tratamento com alimentaçãoDocumento37 páginasDislipidemia: o que é, causas e tratamento com alimentaçãoMaryGuerreroAinda não há avaliações
- Ácidos graxos mono e poli-insaturados na dieta e dislipidemia: benefícios e fontesDocumento30 páginasÁcidos graxos mono e poli-insaturados na dieta e dislipidemia: benefícios e fontesEmiliana100% (1)
- Trabalho LIPIDOSDocumento12 páginasTrabalho LIPIDOSDenilson MiguelAinda não há avaliações
- LipídeosDocumento4 páginasLipídeosrodrigo da costa cardosoAinda não há avaliações
- Atividade 2 - Química e LipídeosDocumento9 páginasAtividade 2 - Química e LipídeosLUIZA NAZARETHAinda não há avaliações
- Dislipidemias e Aterosclerose: Aspectos GeraisDocumento74 páginasDislipidemias e Aterosclerose: Aspectos Geraispatricia torkaski100% (1)
- LIPÍDIOS Fechamento UC IVDocumento8 páginasLIPÍDIOS Fechamento UC IVLuis Mateus Camelo França MartinsAinda não há avaliações
- Aula Cardiovascular AnatomiaDocumento41 páginasAula Cardiovascular AnatomiaAdarly KrothAinda não há avaliações
- Sistema Nervoso PeriféricoDocumento30 páginasSistema Nervoso PeriféricoAdarly KrothAinda não há avaliações
- Aula Cardiovascular - EvoluçãoDocumento15 páginasAula Cardiovascular - EvoluçãoAdarly KrothAinda não há avaliações
- SistRespiratórioDocumento53 páginasSistRespiratórioAdarly KrothAinda não há avaliações
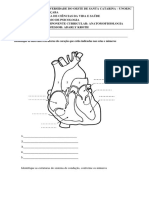
- Estruturas do coração, sistema de condução e respiratórioDocumento2 páginasEstruturas do coração, sistema de condução e respiratórioAdarly KrothAinda não há avaliações
- Sistema Nervoso PeriféricoDocumento30 páginasSistema Nervoso PeriféricoAdarly KrothAinda não há avaliações
- Resenha do livro Novas maneiras de ensinar novas formas de aprenderDocumento3 páginasResenha do livro Novas maneiras de ensinar novas formas de aprenderFelipeMathiasAinda não há avaliações
- História da Tabela PeriódicaDocumento6 páginasHistória da Tabela Periódicaadenilson dos santosAinda não há avaliações
- Sonhos no Terceiro Reich revelam impacto do regime nazistaDocumento17 páginasSonhos no Terceiro Reich revelam impacto do regime nazistaMarcello BecreiAinda não há avaliações
- PORTIFÓLIO - MARILDA GestãoDocumento3 páginasPORTIFÓLIO - MARILDA GestãoMarilia GarciaAinda não há avaliações
- AVANÇADO TREINO 4xDocumento3 páginasAVANÇADO TREINO 4xRaquel SantosAinda não há avaliações
- Modelo Carta Anuencia RSMGDocumento2 páginasModelo Carta Anuencia RSMGbidacasartelAinda não há avaliações
- Visual Novel evoluçãoDocumento10 páginasVisual Novel evoluçãoLincolnAinda não há avaliações
- Avaliação - 2º Tri - Cadernos de QuestõesDocumento9 páginasAvaliação - 2º Tri - Cadernos de QuestõesLucas RodriguesAinda não há avaliações
- Dinâmicas para integração no 1o diaDocumento2 páginasDinâmicas para integração no 1o diaNoemi NonatoAinda não há avaliações
- Orçamento ParticipativoDocumento13 páginasOrçamento ParticipativoIvone Pedroso100% (1)
- O modelo francês de fomento ao cinemaDocumento10 páginasO modelo francês de fomento ao cinemaGil VicenteAinda não há avaliações
- Manual TambaquiDocumento48 páginasManual TambaquiIsaque Leal100% (1)
- Introdução Andebol FranciscoDocumento10 páginasIntrodução Andebol FranciscoAnabela Gutierres CunhaAinda não há avaliações
- Poder Executivo: Iário FicialDocumento152 páginasPoder Executivo: Iário FicialRaul Tavares CecattoAinda não há avaliações
- Frase de desculpas: 6 elementos e 7 formasDocumento4 páginasFrase de desculpas: 6 elementos e 7 formasGabrielle RangelAinda não há avaliações
- Origem Do Eletromagnetismo e Física ModernaDocumento44 páginasOrigem Do Eletromagnetismo e Física ModernaCleniwton AlvesAinda não há avaliações
- Aula 1 - Pei Ii 1.2 - 2022.2Documento18 páginasAula 1 - Pei Ii 1.2 - 2022.2harlley sanAinda não há avaliações
- Processo de Compostagem de Resíduos Urbanos em PequenaDocumento124 páginasProcesso de Compostagem de Resíduos Urbanos em PequenaMIGAJOHNSONAinda não há avaliações
- Ficha Tecnica 15 Shiver Bass 3.8 4 Ohms A4Documento2 páginasFicha Tecnica 15 Shiver Bass 3.8 4 Ohms A4wendlyAinda não há avaliações
- Apontamentos de EDIFICIOS - Cap. 1 - CONCEPCAO ESTRUTURAL DE EDIFICIOS. ANALISE ESTRUTURAL RevAGO2019Documento47 páginasApontamentos de EDIFICIOS - Cap. 1 - CONCEPCAO ESTRUTURAL DE EDIFICIOS. ANALISE ESTRUTURAL RevAGO2019Arilson Ngoca100% (1)
- TESTE de Atividade ATEXDocumento3 páginasTESTE de Atividade ATEXAndreia SilvaAinda não há avaliações
- Dimensionamento da rede elétrica para MungassaDocumento113 páginasDimensionamento da rede elétrica para MungassachalepedroAinda não há avaliações
- Arduino, Engenharia e TIDocumento35 páginasArduino, Engenharia e TIFrancisco AlbertoAinda não há avaliações
- Bircat Hamazon - Judaísmo Na PráticaDocumento9 páginasBircat Hamazon - Judaísmo Na PráticaMarco PoleseAinda não há avaliações
- Cristais de Cura e ProtecaoDocumento8 páginasCristais de Cura e ProtecaoanagrazielaAinda não há avaliações
- Diretrizes Brasileiras para o - Diagnóstico de Fibrose CísticaDocumento27 páginasDiretrizes Brasileiras para o - Diagnóstico de Fibrose CísticaJosé Elias BarrosAinda não há avaliações
- Escolha múltipla sobre forças gravitacionaisDocumento1 páginaEscolha múltipla sobre forças gravitacionaissergiopcm8997Ainda não há avaliações
- PDCADocumento4 páginasPDCARoger Paulo Barbosa ChavesAinda não há avaliações
- A pequena vendedora de fósforosDocumento4 páginasA pequena vendedora de fósforosTiago dos AnjosAinda não há avaliações
- Questionário I - Diversidade Linguística e ComunicaçãoDocumento3 páginasQuestionário I - Diversidade Linguística e ComunicaçãoAnalice CabralAinda não há avaliações
- DIETA DASH PARA INICIANTES: Obtenha uma vida saudável perdendo peso de forma rápida, saudável e equilibrada. Reduzir a pressão arterial, inchaço abdominal e inflamação corporalNo EverandDIETA DASH PARA INICIANTES: Obtenha uma vida saudável perdendo peso de forma rápida, saudável e equilibrada. Reduzir a pressão arterial, inchaço abdominal e inflamação corporalNota: 4 de 5 estrelas4/5 (14)
- Detox de 10 dias: Como os sucos verdes limpam o seu organismo e emagrecemNo EverandDetox de 10 dias: Como os sucos verdes limpam o seu organismo e emagrecemNota: 2 de 5 estrelas2/5 (1)
- Sem glúten: Pães, tortas e bolos sem restriçõesNo EverandSem glúten: Pães, tortas e bolos sem restriçõesNota: 5 de 5 estrelas5/5 (3)
- Hipertrofia Muscular: da Nutrição à SuplementaçãoNo EverandHipertrofia Muscular: da Nutrição à SuplementaçãoAinda não há avaliações
- Uma dieta além da moda: Uma abordagem científica para a perda de peso e a manutenção da sáudeNo EverandUma dieta além da moda: Uma abordagem científica para a perda de peso e a manutenção da sáudeNota: 4.5 de 5 estrelas4.5/5 (4)
- Dieta Para Gordura no Fígado Em português/ Diet For Fat In The Liver In PortugueseNo EverandDieta Para Gordura no Fígado Em português/ Diet For Fat In The Liver In PortugueseNota: 5 de 5 estrelas5/5 (1)
- Saúde Intestinal, Limpeza Hepática, Fadiga Adrenal, Depressão, Ansiedade, Perda De Peso E Pele Sadia: Cure Sua Mente, Corpo E Hormônios Com Dieta, Nutrição, Medicina Natural E Terapias HolísticasNo EverandSaúde Intestinal, Limpeza Hepática, Fadiga Adrenal, Depressão, Ansiedade, Perda De Peso E Pele Sadia: Cure Sua Mente, Corpo E Hormônios Com Dieta, Nutrição, Medicina Natural E Terapias HolísticasNota: 5 de 5 estrelas5/5 (7)
- Dieta Anti-inflamatória EstratégicaNo EverandDieta Anti-inflamatória EstratégicaNota: 5 de 5 estrelas5/5 (2)
- Livro de Receitas da Dieta Anti-inflamatória de 7 Dias E Plano Fácil de Reduzir a Inflamação Em portuguêsNo EverandLivro de Receitas da Dieta Anti-inflamatória de 7 Dias E Plano Fácil de Reduzir a Inflamação Em portuguêsNota: 5 de 5 estrelas5/5 (4)
- 21 HiperSucos: para saúde física, mental e longevidadeNo Everand21 HiperSucos: para saúde física, mental e longevidadeAinda não há avaliações
- Este não é mais um livro de dieta: O novo e libertador estilo de vida alimentar para saúde e boa forma que derruba o conceito de dietasNo EverandEste não é mais um livro de dieta: O novo e libertador estilo de vida alimentar para saúde e boa forma que derruba o conceito de dietasNota: 4.5 de 5 estrelas4.5/5 (30)
- Cozinha Vegana: 80 Receitas Práticas, Saudáveis E SaborosasNo EverandCozinha Vegana: 80 Receitas Práticas, Saudáveis E SaborosasAinda não há avaliações
- A alquimia dos sabores: a culinária funcional: Edição revista e atualizadaNo EverandA alquimia dos sabores: a culinária funcional: Edição revista e atualizadaAinda não há avaliações
- Dieta Mediterrânea para Iniciantes Em português/ Mediterranean Diet for Beginners In PortugueseNo EverandDieta Mediterrânea para Iniciantes Em português/ Mediterranean Diet for Beginners In PortugueseAinda não há avaliações
- Treino de Peso Corporal Em português/ Body Weight Training In PortugueseNo EverandTreino de Peso Corporal Em português/ Body Weight Training In PortugueseAinda não há avaliações
- Mais energia: Dieta para aumentar a resistência e dar mais energiaNo EverandMais energia: Dieta para aumentar a resistência e dar mais energiaAinda não há avaliações
- Mente sã, corpo são: O poder de uma mente saudávelNo EverandMente sã, corpo são: O poder de uma mente saudávelNota: 5 de 5 estrelas5/5 (3)
- Abandonando o Padrão da Obesidade através da Constelação SistêmicaNo EverandAbandonando o Padrão da Obesidade através da Constelação SistêmicaNota: 5 de 5 estrelas5/5 (5)
- Dieta Mediterrânea - A Ciência E A Arte Da Dieta Mediterrânea: Um Guia Completo Para Iniciantes Para Queimar Gordura E Alcançar Uma Perda De Peso PermanenteNo EverandDieta Mediterrânea - A Ciência E A Arte Da Dieta Mediterrânea: Um Guia Completo Para Iniciantes Para Queimar Gordura E Alcançar Uma Perda De Peso PermanenteAinda não há avaliações
- Pequenos Hábitos Para A Perda De Peso: Esqueça As Dietas Keto, Paleo, Mediterrânea Ou Vegetariana: Construa Facilmente Novos Hábitos Saudáveis Sem Sofrer - Domine O Jogo Interno Da SaúdeNo EverandPequenos Hábitos Para A Perda De Peso: Esqueça As Dietas Keto, Paleo, Mediterrânea Ou Vegetariana: Construa Facilmente Novos Hábitos Saudáveis Sem Sofrer - Domine O Jogo Interno Da SaúdeAinda não há avaliações
- O Grande Livro Da Dieta Cetogênica CulináriaNo EverandO Grande Livro Da Dieta Cetogênica CulináriaAinda não há avaliações