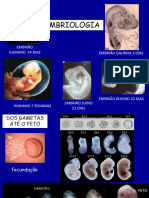
Você também pode gostar
- Reprodução e cuidados com potros equinosDocumento6 páginasReprodução e cuidados com potros equinosAna ZoccolerAinda não há avaliações
- DEFINIÇÃO_DE_ALGUNS_CONCEITOS_2015Documento34 páginasDEFINIÇÃO_DE_ALGUNS_CONCEITOS_2015Júnior SilvérioAinda não há avaliações
- Fecundação e formação do zigotoDocumento10 páginasFecundação e formação do zigotoRhosula sandesAinda não há avaliações
- Fecundação e DesenvolvimentoDocumento7 páginasFecundação e DesenvolvimentoFernando A VilelaAinda não há avaliações
- Bases da reprodução humanaDocumento18 páginasBases da reprodução humanaMaria LuciaAinda não há avaliações
- EmbriologiaDocumento21 páginasEmbriologiaGilberto Cavalheiro VieiraAinda não há avaliações
- Ovulação, Fertilização e Gestação dos animaisDocumento15 páginasOvulação, Fertilização e Gestação dos animaisCalisto MelhoAinda não há avaliações
- Veterinarian Docs ObstetriciaDocumento42 páginasVeterinarian Docs ObstetriciaDácia ArrudaAinda não há avaliações
- Susana 065329Documento14 páginasSusana 065329Braiton antonioAinda não há avaliações
- Desenvolvimento pré-natal e fatores da personalidadeDocumento12 páginasDesenvolvimento pré-natal e fatores da personalidadePaulo De CristoAinda não há avaliações
- Anatomia e fisiologia do trato reprodutivo femininoDocumento46 páginasAnatomia e fisiologia do trato reprodutivo femininoBruna Milagre80% (5)
- Processo de fecundação e desenvolvimento embrionárioDocumento12 páginasProcesso de fecundação e desenvolvimento embrionárioGabriela RochaAinda não há avaliações
- Tutoria Problema I - Fechamento Ii 1. Descrever As Fases Da FecundaçãoDocumento12 páginasTutoria Problema I - Fechamento Ii 1. Descrever As Fases Da FecundaçãoBárbara Cristal Oliveira AlmeidaAinda não há avaliações
- Embriologia - AULA 03Documento60 páginasEmbriologia - AULA 03Emanuelly MatosAinda não há avaliações
- Resumo de Obstetrícia - Parte IDocumento21 páginasResumo de Obstetrícia - Parte IHernane Ah100% (1)
- Trabalho de Histologia - Estudo DirigidoDocumento7 páginasTrabalho de Histologia - Estudo DirigidoThais Lorrany K. AntunesAinda não há avaliações
- Desenvolvimento EmbrionarioDocumento10 páginasDesenvolvimento Embrionariogeronim AlbertAinda não há avaliações
- Embriologia - Caderno e ResumoDocumento26 páginasEmbriologia - Caderno e ResumoJôsymércio Farias100% (1)
- Fisiologia do Sistema ReprodutorDocumento7 páginasFisiologia do Sistema ReprodutorMatheus R10Ainda não há avaliações
- Fisiologia Reprodutiva Das Aves - 2013Documento9 páginasFisiologia Reprodutiva Das Aves - 2013Thiago MendzAinda não há avaliações
- Histologia e EmbriologiaDocumento45 páginasHistologia e Embriologiaadrienerosceli100% (3)
- 50 QuestõesDocumento9 páginas50 QuestõesFernanda MrsAinda não há avaliações
- Embriologia Humana: Fases da FertilizaçãoDocumento8 páginasEmbriologia Humana: Fases da FertilizaçãoNascimento Jga100% (1)
- 3 FecundaçãoDocumento6 páginas3 FecundaçãoIhury JhonsonAinda não há avaliações
- Reprodução em CefalópodesDocumento22 páginasReprodução em CefalópodesIsabela CabralAinda não há avaliações
- Primeira semana de gestaçãoDocumento21 páginasPrimeira semana de gestaçãoJoao V LenzAinda não há avaliações
- Fisiologia AnimalDocumento11 páginasFisiologia AnimalERIKA APARECIDA SILVA NAVASAinda não há avaliações
- gestaçãoDocumento17 páginasgestaçãofernandalacenAinda não há avaliações
- Observação de Cortes Histológicos de Testículo e Ovário de CoelhosDocumento11 páginasObservação de Cortes Histológicos de Testículo e Ovário de CoelhosBárbara LaranjoAinda não há avaliações
- Capitulo 1 Vitelogenese Nos Ecdizoa - Nematoides e Insetos Como Exemplos PDFDocumento45 páginasCapitulo 1 Vitelogenese Nos Ecdizoa - Nematoides e Insetos Como Exemplos PDFJose Carlos Pelielo MattosAinda não há avaliações
- Embriologia Humana - Fecundação, Clivagem, Compactação, Mórula, Blástula e Nidação, Gastrulação, Fechamento Do Embrião, Noções de Organogenese e TeratogeneseDocumento20 páginasEmbriologia Humana - Fecundação, Clivagem, Compactação, Mórula, Blástula e Nidação, Gastrulação, Fechamento Do Embrião, Noções de Organogenese e TeratogeneseAmanda AraujoAinda não há avaliações
- Questionario Av2 2021Documento5 páginasQuestionario Av2 2021Letycia Fernandes100% (1)
- Ovários, ciclo estral e reprodução nas fêmeasDocumento15 páginasOvários, ciclo estral e reprodução nas fêmeasWerner AlbuquerqueAinda não há avaliações
- Anatomofisiologia Comparada Laboratório 5Documento6 páginasAnatomofisiologia Comparada Laboratório 5Adilson JuniorAinda não há avaliações
- Desenvolvimento embrionário humano desde a fecundação até a gastrulaçãoDocumento8 páginasDesenvolvimento embrionário humano desde a fecundação até a gastrulaçãoJhon ChagasAinda não há avaliações
- Resolucao Dos Exercicios Do Manual Biodesafios 12o AnoDocumento86 páginasResolucao Dos Exercicios Do Manual Biodesafios 12o AnoBeatriz ArnaudAinda não há avaliações
- Parte 1 FemeaDocumento71 páginasParte 1 Femeadanielleferreirat2Ainda não há avaliações
- Gametogenese 2020 MEDVETDocumento72 páginasGametogenese 2020 MEDVETMaisa CeciliaAinda não há avaliações
- Gestação, Parto e LactaçãoDocumento17 páginasGestação, Parto e LactaçãoReginaAinda não há avaliações
- Fecundação RESUMODocumento6 páginasFecundação RESUMOana filipeAinda não há avaliações
- Aula 1 Fecundacao PDF UnicoDocumento76 páginasAula 1 Fecundacao PDF UniconmanoelrogerAinda não há avaliações
- Aula 02 Obstetricia - Da Fecundação Ao DesenvolvimentoDocumento53 páginasAula 02 Obstetricia - Da Fecundação Ao DesenvolvimentoAna CamilaAinda não há avaliações
- Funções reprodutivas e hormonais masculinasDocumento9 páginasFunções reprodutivas e hormonais masculinasRicardo Gomes Fernandes0% (1)
- Transporte de Gametas e FertilizaçãoDocumento4 páginasTransporte de Gametas e FertilizaçãoRafael de Lima SantosAinda não há avaliações
- Embriologia Animal: Gametogênese, Fecundação e ImplantaçãoDocumento23 páginasEmbriologia Animal: Gametogênese, Fecundação e ImplantaçãoCarolina Araújo100% (1)
- Gametogênese e FertilizaçãoDocumento38 páginasGametogênese e FertilizaçãoLarissa AlvesAinda não há avaliações
- Rel Bio 12Documento6 páginasRel Bio 12Luísa Braz TeixeiraAinda não há avaliações
- Anatomia e Fisiologia Do Cilco Estral de Éguas e Vacas (Salvo Automaticamente) - 1Documento42 páginasAnatomia e Fisiologia Do Cilco Estral de Éguas e Vacas (Salvo Automaticamente) - 1Gleison JustinoAinda não há avaliações
- Morfofisiologia MasculinoDocumento8 páginasMorfofisiologia MasculinoAdolfo SilvaAinda não há avaliações
- Biologia Do Desenvolvimento de MamíferosDocumento11 páginasBiologia Do Desenvolvimento de MamíferosGabi SoaresAinda não há avaliações
- Desenvolvimento embrionário de répteisDocumento4 páginasDesenvolvimento embrionário de répteisLiziane RodriguesAinda não há avaliações
- Biologia 12º Ano ReproduçãoDocumento10 páginasBiologia 12º Ano ReproduçãoRaquelAinda não há avaliações
- Reproducao AvesDocumento11 páginasReproducao AvesbiancadrumondAinda não há avaliações
- Escopo Anatomia HumanaDocumento6 páginasEscopo Anatomia HumanaRosana LimaAinda não há avaliações
- Anatomia reprodutiva masculina dos mamíferos domésticosDocumento14 páginasAnatomia reprodutiva masculina dos mamíferos domésticosYasmin SaetherAinda não há avaliações
- Sistema reprodutor e genética humanosDocumento6 páginasSistema reprodutor e genética humanosTeresa BorgesAinda não há avaliações
- GravidezAdolescenteDocumento51 páginasGravidezAdolescenteLara JessicaAinda não há avaliações
- As particularidades da gestação caninaDocumento16 páginasAs particularidades da gestação caninaRachel GreenAinda não há avaliações
- Caderno PRESSE 3º Ciclo 2014Documento110 páginasCaderno PRESSE 3º Ciclo 2014luper4609Ainda não há avaliações
- Teste Ciencias 9 Ano Reproducao Genetica e FecundacaoDocumento8 páginasTeste Ciencias 9 Ano Reproducao Genetica e FecundacaosabercommeritoAinda não há avaliações
- Bases anatômicas e fisiológicas da sexualidade humana: Fisiologia do coitoDocumento154 páginasBases anatômicas e fisiológicas da sexualidade humana: Fisiologia do coitoCristina SilvaAinda não há avaliações
- MilhoDocumento333 páginasMilhoEdmar TuelherAinda não há avaliações
- Ciencias Natureza Nova Eja Aluno Mod02 Vol01 PDFDocumento424 páginasCiencias Natureza Nova Eja Aluno Mod02 Vol01 PDFPriscila MonteiroAinda não há avaliações
- DESENVOLVIMENTO HUMANODocumento6 páginasDESENVOLVIMENTO HUMANOPaulo RodrigoAinda não há avaliações
- 2019-11!20!11!51!17dissertação - Mariana Da Mata SilveiraDocumento60 páginas2019-11!20!11!51!17dissertação - Mariana Da Mata SilveiraPedro LuizAinda não há avaliações
- 3a Aula AULA Efeito Materno 23Documento89 páginas3a Aula AULA Efeito Materno 23João GarciaAinda não há avaliações
- Espiritismo e AbortoDocumento105 páginasEspiritismo e AbortoJorge ArieiroAinda não há avaliações
- Citologia - Células DeciduaisDocumento2 páginasCitologia - Células DeciduaisAline GrazieleAinda não há avaliações
- Rehagro - Artigos Técnicos - Inseminação Artificial, Monta Natural e Monta Natural Controlada - Vantagens e DesvantagensDocumento5 páginasRehagro - Artigos Técnicos - Inseminação Artificial, Monta Natural e Monta Natural Controlada - Vantagens e DesvantagensGislaine CostaAinda não há avaliações
- Ficha de Reforço 9 A Reprodução Nos AnimaisDocumento2 páginasFicha de Reforço 9 A Reprodução Nos AnimaisSusana AndradeAinda não há avaliações
- FPS Medicina 2018.1Documento99 páginasFPS Medicina 2018.1liviaAinda não há avaliações
- Apostila Fisiologia Vegetal FrutificaçãoDocumento13 páginasApostila Fisiologia Vegetal FrutificaçãofrancohoffAinda não há avaliações
- O conto de fadas de SuriáDocumento12 páginasO conto de fadas de SuriáAlessandra PancieriAinda não há avaliações
- TeriogenologiaDocumento7 páginasTeriogenologiaFilipe MartinsAinda não há avaliações
- Boletim Técnico N 136: Manual de Reprodução de Peixes de Água Doce Com Cultivo Comercial Na Região Sul Do BrasilDocumento55 páginasBoletim Técnico N 136: Manual de Reprodução de Peixes de Água Doce Com Cultivo Comercial Na Região Sul Do BrasilclaudiousdecastroAinda não há avaliações
- Prova Faresi 2023-2-15 FinalDocumento29 páginasProva Faresi 2023-2-15 Final8gmt2vtkrqAinda não há avaliações
- Gabarito - Lista ReproduçãoDocumento2 páginasGabarito - Lista Reproduçãopajexo4126Ainda não há avaliações
- Ciclo de vida de projetos: entenda as 4 fasesDocumento5 páginasCiclo de vida de projetos: entenda as 4 fasesdiogo Barbosa De AlencarAinda não há avaliações
- Atlas Morfologia1412 PDFDocumento99 páginasAtlas Morfologia1412 PDFKaren OezauAinda não há avaliações
- Duração sangramento nidaçãoDocumento1 páginaDuração sangramento nidaçãoFernando LêdoAinda não há avaliações
- Atividades Geral 5 Ano IIDocumento5 páginasAtividades Geral 5 Ano IISirlene MazottiAinda não há avaliações
- Resoluo N. 2294 2021 Do CFMDocumento8 páginasResoluo N. 2294 2021 Do CFMRégis lewsAinda não há avaliações
- Como Orar Pela Cura Interior Pelas Fases Da VidaDocumento5 páginasComo Orar Pela Cura Interior Pelas Fases Da VidaJhonatan Machado100% (2)
- Teorias Psicológicas Do DesenvolvimentoDocumento44 páginasTeorias Psicológicas Do DesenvolvimentovkarmoAinda não há avaliações
- Exercícios Reprodução 3º AnoDocumento4 páginasExercícios Reprodução 3º AnoBeatriz De limaAinda não há avaliações
- Transferencia Intratubaria de GametasDocumento8 páginasTransferencia Intratubaria de GametassoparaalgumascoisasAinda não há avaliações
- Treinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNo EverandTreinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNota: 4.5 de 5 estrelas4.5/5 (169)
- Biomecânica Básica dos Exercícios: Membros InferioresNo EverandBiomecânica Básica dos Exercícios: Membros InferioresNota: 3.5 de 5 estrelas3.5/5 (8)
- Uma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNo EverandUma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNota: 4 de 5 estrelas4/5 (13)
- Atlas de anatomia: Imagens humanas e veterináriasNo EverandAtlas de anatomia: Imagens humanas e veterináriasNota: 4.5 de 5 estrelas4.5/5 (2)
- Serviços Ecossistêmicos e Planejamento Urbano: A Natureza a Favor do Desenvolvimento Sustentável das CidadesNo EverandServiços Ecossistêmicos e Planejamento Urbano: A Natureza a Favor do Desenvolvimento Sustentável das CidadesAinda não há avaliações
- Parasitologia 1: Helmintos de Interesse MédicoNo EverandParasitologia 1: Helmintos de Interesse MédicoAinda não há avaliações
- Educação ambiental: A formação do sujeito ecológicoNo EverandEducação ambiental: A formação do sujeito ecológicoAinda não há avaliações
- Equilíbrio hormonal _ Recupere equilíbrio hormonal, libido, sono e emagreça já!No EverandEquilíbrio hormonal _ Recupere equilíbrio hormonal, libido, sono e emagreça já!Nota: 5 de 5 estrelas5/5 (2)
- Fundamentos Em Cirurgia OralNo EverandFundamentos Em Cirurgia OralNota: 5 de 5 estrelas5/5 (1)
- Psicoterapia: Teorias e técnicas psicoterápicasNo EverandPsicoterapia: Teorias e técnicas psicoterápicasNota: 5 de 5 estrelas5/5 (4)
- Suplementação proteica: Aminoacidemia e saúde vascularNo EverandSuplementação proteica: Aminoacidemia e saúde vascularAinda não há avaliações
- A clínica gestáltica com adolescentes: Caminhos clínicos e institucionaisNo EverandA clínica gestáltica com adolescentes: Caminhos clínicos e institucionaisNota: 3.5 de 5 estrelas3.5/5 (3)
- Fisiologia Cardiorrespiratória Ensino SuperiorNo EverandFisiologia Cardiorrespiratória Ensino SuperiorAinda não há avaliações
- Topônimos no Sul da Bahia: nominações dos municípios originados da capitania hereditária de São Jorge dos Ilheus (1950 a 1960)No EverandTopônimos no Sul da Bahia: nominações dos municípios originados da capitania hereditária de São Jorge dos Ilheus (1950 a 1960)Nota: 5 de 5 estrelas5/5 (1)
- Concentração: Torne-se mais consciente e economize tempoNo EverandConcentração: Torne-se mais consciente e economize tempoNota: 4.5 de 5 estrelas4.5/5 (3)