Você também pode gostar
- 3 FecundaçãoDocumento6 páginas3 FecundaçãoIhury JhonsonAinda não há avaliações
- Histologia e EmbriologiaDocumento45 páginasHistologia e Embriologiaadrienerosceli100% (3)
- PDF Landim Biologia - Parte 28Documento3 páginasPDF Landim Biologia - Parte 28João GabrielAinda não há avaliações
- Embriologia Humana: Fases da FertilizaçãoDocumento8 páginasEmbriologia Humana: Fases da FertilizaçãoNascimento Jga100% (1)
- Fisioreprobovina 3 Gestao 1Documento20 páginasFisioreprobovina 3 Gestao 1vinicius.maretto25Ainda não há avaliações
- Biologia 2 - Reprodução Humana, Embriologia, Histologia AnimalDocumento633 páginasBiologia 2 - Reprodução Humana, Embriologia, Histologia Animal《B4》Klaus'zAinda não há avaliações
- Processo de fecundação e desenvolvimento embrionárioDocumento12 páginasProcesso de fecundação e desenvolvimento embrionárioGabriela RochaAinda não há avaliações
- Desenvolvimento embrionário de répteisDocumento4 páginasDesenvolvimento embrionário de répteisLiziane RodriguesAinda não há avaliações
- ESTUDO DIRIGIDO 4 - EmbriologiaDocumento4 páginasESTUDO DIRIGIDO 4 - EmbriologiaCarla Caroline Alves De LimaAinda não há avaliações
- GametogêneseDocumento2 páginasGametogêneseKarineAinda não há avaliações
- Fecundação: início do desenvolvimento humanoDocumento3 páginasFecundação: início do desenvolvimento humanoRoberth Alexandro Soares de OliveiraAinda não há avaliações
- Fecundação e DesenvolvimentoDocumento7 páginasFecundação e DesenvolvimentoFernando A VilelaAinda não há avaliações
- Anatomofisiologia Comparada Laboratório 5Documento6 páginasAnatomofisiologia Comparada Laboratório 5Adilson JuniorAinda não há avaliações
- Reprodução Humana-FecundaçãoDocumento4 páginasReprodução Humana-FecundaçãoBeatrizAinda não há avaliações
- Tutoria Problema I - Fechamento Ii 1. Descrever As Fases Da FecundaçãoDocumento12 páginasTutoria Problema I - Fechamento Ii 1. Descrever As Fases Da FecundaçãoBárbara Cristal Oliveira AlmeidaAinda não há avaliações
- Preparando Gametas e Iniciando Os Eventos EmbrionáriosDocumento15 páginasPreparando Gametas e Iniciando Os Eventos EmbrionáriosalcantarabrennaAinda não há avaliações
- Questionario Av2 2021Documento5 páginasQuestionario Av2 2021Letycia Fernandes100% (1)
- Fertilização - Aula 5Documento23 páginasFertilização - Aula 5Gabriel Henrique LopesAinda não há avaliações
- 4-1 Aula BiocelDocumento51 páginas4-1 Aula BiocelFelipeMarodinAinda não há avaliações
- Primeira Semana Desenvolvimento EmbrionárioDocumento11 páginasPrimeira Semana Desenvolvimento EmbrionárioLone GhostAinda não há avaliações
- Biologia 1 - GametogêneseDocumento11 páginasBiologia 1 - GametogêneseVIVI LOUSANOAinda não há avaliações
- Reproducao - Gametogenese e FecundaçãoDocumento73 páginasReproducao - Gametogenese e FecundaçãoKerle1100% (2)
- Embriologia Humana - Fecundação, Clivagem, Compactação, Mórula, Blástula e Nidação, Gastrulação, Fechamento Do Embrião, Noções de Organogenese e TeratogeneseDocumento20 páginasEmbriologia Humana - Fecundação, Clivagem, Compactação, Mórula, Blástula e Nidação, Gastrulação, Fechamento Do Embrião, Noções de Organogenese e TeratogeneseAmanda AraujoAinda não há avaliações
- Questões - MicologiaDocumento6 páginasQuestões - MicologiaJadson NascimentoAinda não há avaliações
- Estudo Dirigido Embrio IDocumento4 páginasEstudo Dirigido Embrio IdaienescamposAinda não há avaliações
- Reprodução em CefalópodesDocumento22 páginasReprodução em CefalópodesIsabela CabralAinda não há avaliações
- Mecanismos ReprodutivosDocumento31 páginasMecanismos ReprodutivosGiovana Raquel Pagelkopf100% (1)
- Embriologia - Caderno e ResumoDocumento26 páginasEmbriologia - Caderno e ResumoJôsymércio Farias100% (1)
- Biologia 12.º AnoDocumento65 páginasBiologia 12.º AnoPaula Amado100% (1)
- Sist Rep HumanoDocumento40 páginasSist Rep Humanojoaopedroolk2129Ainda não há avaliações
- M7P2 - Embriogênese, teratogenia e abortamentoDocumento16 páginasM7P2 - Embriogênese, teratogenia e abortamentoGustavo AraújoAinda não há avaliações
- FertilizaçãoDocumento3 páginasFertilizaçãostd medAinda não há avaliações
- Fecundação, Desenvolvimento Emb.e PartoDocumento59 páginasFecundação, Desenvolvimento Emb.e PartoRita RodriguesAinda não há avaliações
- Sistemas Reprodutores AnimaisDocumento8 páginasSistemas Reprodutores AnimaisJason Pereira100% (1)
- Apresentava Sobre Formação Do EmbriãoDocumento10 páginasApresentava Sobre Formação Do EmbriãoelytopedrooAinda não há avaliações
- Genética e Embriologia - Fecundação e ClivagemDocumento13 páginasGenética e Embriologia - Fecundação e ClivagemJoice CruzAinda não há avaliações
- Relatório 9Documento2 páginasRelatório 9martim.m.ezequielAinda não há avaliações
- EmbriologiaDocumento8 páginasEmbriologiaThiago Soldatelli BritoAinda não há avaliações
- Embriologia - AULA 03Documento60 páginasEmbriologia - AULA 03Emanuelly MatosAinda não há avaliações
- Gametogénese e Regulação HormonalDocumento53 páginasGametogénese e Regulação HormonalMaria SantosAinda não há avaliações
- Fecundação e formação do zigotoDocumento10 páginasFecundação e formação do zigotoRhosula sandesAinda não há avaliações
- Bio ReproducaoDocumento29 páginasBio ReproducaoLeandra SabinoAinda não há avaliações
- 12º BiologiaDocumento12 páginas12º BiologiaMiriam CruzAinda não há avaliações
- Primeira semana de gestaçãoDocumento21 páginasPrimeira semana de gestaçãoJoao V LenzAinda não há avaliações
- 3 - GAMETOGENESE - FECUNDAÇÃO (Salvo Automaticamente)Documento37 páginas3 - GAMETOGENESE - FECUNDAÇÃO (Salvo Automaticamente)Magno SilvaAinda não há avaliações
- Processo de Fecundação emDocumento29 páginasProcesso de Fecundação emPaulo Henrique SoaresAinda não há avaliações
- Reprodução Sexuada 1Documento9 páginasReprodução Sexuada 1Jeesiel TemoteoAinda não há avaliações
- Aula Dra Ana PaulaDocumento4 páginasAula Dra Ana PaulaLêh GomesAinda não há avaliações
- Embriologia e reprodução marinhaDocumento5 páginasEmbriologia e reprodução marinhaLIDIA PUGLIESSAAinda não há avaliações
- Gametogênese HumanaDocumento43 páginasGametogênese HumanaLucas GomesAinda não há avaliações
- Desenvolvimento pré-natal e fatores da personalidadeDocumento12 páginasDesenvolvimento pré-natal e fatores da personalidadePaulo De CristoAinda não há avaliações
- Biologia Do Desenvolvimento de MamíferosDocumento11 páginasBiologia Do Desenvolvimento de MamíferosGabi SoaresAinda não há avaliações
- Fertilização e Clivagem PDFDocumento69 páginasFertilização e Clivagem PDFGabrielleGesserAinda não há avaliações
- Resumo de Embriologia PDFDocumento17 páginasResumo de Embriologia PDFDanielaAlvesAinda não há avaliações
- Fertilização, clivagem e implantação inicialDocumento8 páginasFertilização, clivagem e implantação inicialMaria Esthela Bull LooseAinda não há avaliações
- Exercícios de Fixação Ovogênese - QUASEDocumento6 páginasExercícios de Fixação Ovogênese - QUASEGiovanni Ataíde100% (1)
- Biologia TeteDocumento10 páginasBiologia TeteBaptista ZaqueuAinda não há avaliações
- Apostila Metodo AprovacaoDocumento20 páginasApostila Metodo AprovacaoGilberto Cavalheiro VieiraAinda não há avaliações
- + DE 100 Agro Mapas Mentais Antes Da PorteiraDocumento127 páginas+ DE 100 Agro Mapas Mentais Antes Da PorteiraDaniel MocellinAinda não há avaliações
- Aula AvesDocumento39 páginasAula AvesGilberto Cavalheiro Vieira100% (1)
- A Teoria de Resposta Ao ItemDocumento4 páginasA Teoria de Resposta Ao ItemGilberto Cavalheiro VieiraAinda não há avaliações
- Diferença Entre Peixes de Água Salgada e Peixes e Água DoceDocumento12 páginasDiferença Entre Peixes de Água Salgada e Peixes e Água DoceGilberto Cavalheiro VieiraAinda não há avaliações
- Curso de Inverno de Biologia Evolutiva - Evo-Devo e Epigenética (Gilberto Cavalheiro Vieira)Documento21 páginasCurso de Inverno de Biologia Evolutiva - Evo-Devo e Epigenética (Gilberto Cavalheiro Vieira)Gilberto Cavalheiro VieiraAinda não há avaliações
- Aula CitologiaDocumento25 páginasAula CitologiaGilberto Cavalheiro VieiraAinda não há avaliações
- A Pres en Tao 1Documento14 páginasA Pres en Tao 1Gilberto Cavalheiro VieiraAinda não há avaliações
- Aula AngiospermasDocumento6 páginasAula AngiospermasGilberto Cavalheiro VieiraAinda não há avaliações
- Ficha Nº 4 O NomeDocumento9 páginasFicha Nº 4 O NomeFilipa DraiblateAinda não há avaliações
- Exercicio de CircuitosDocumento2 páginasExercicio de CircuitosFrancisco SilvaAinda não há avaliações
- Document - Onl - o Maior Segredo de Todos Os Tempos para Criar Dinheiro Joe VitaleDocumento51 páginasDocument - Onl - o Maior Segredo de Todos Os Tempos para Criar Dinheiro Joe VitaleAzevedoAinda não há avaliações
- Frater Achad - 31 Hinos A Deusa Das Estrelas (PT)Documento18 páginasFrater Achad - 31 Hinos A Deusa Das Estrelas (PT)Caléu N. MoraesAinda não há avaliações
- Dízimo Mirim: Ensinando a Prática do Dízimo na CatequeseDocumento8 páginasDízimo Mirim: Ensinando a Prática do Dízimo na CatequesegracyynhaAinda não há avaliações
- Cadeno de Geografia 8ºanoDocumento14 páginasCadeno de Geografia 8ºanoMarlene RicardoAinda não há avaliações
- Resenha Sobre ValsinerDocumento3 páginasResenha Sobre Valsinerveridiana_205804362Ainda não há avaliações
- Catálogo de Acessórios AutomotivosDocumento12 páginasCatálogo de Acessórios AutomotivosNando StremelAinda não há avaliações
- As colunas na Maçonaria: significado simbólicoDocumento7 páginasAs colunas na Maçonaria: significado simbólicoFranciano Vieira Pires100% (1)
- Os recursos hídricos em PortugalDocumento5 páginasOs recursos hídricos em PortugalMargaridaAinda não há avaliações
- NIT Dicap 1 - 03Documento10 páginasNIT Dicap 1 - 03xeseAinda não há avaliações
- Transtorno de LinguagemDocumento8 páginasTranstorno de LinguagemmetalliraphaAinda não há avaliações
- Resumo de Psicofarmacologia da Faculdade Internacional da ParaíbaDocumento6 páginasResumo de Psicofarmacologia da Faculdade Internacional da ParaíbaAphrouddy (Koi Ni Furu)Ainda não há avaliações
- Ficha Avaliação EL - Robinson Crusoe - (Correção)Documento4 páginasFicha Avaliação EL - Robinson Crusoe - (Correção)Patrícia Valente100% (1)
- Manto PeridotíticoDocumento367 páginasManto Peridotítico19-2011ºB Ana Sofia Ferreira DuarteAinda não há avaliações
- Avaliar a caminhada da catequeseDocumento13 páginasAvaliar a caminhada da catequeseJoão MeloAinda não há avaliações
- Artigo - DesignDocumento15 páginasArtigo - DesignFábio Henrique Lucas da CostaAinda não há avaliações
- Relatório parcial de Psicologia ExperimentalDocumento16 páginasRelatório parcial de Psicologia ExperimentalGEOVANA RISSATO GARCIAAinda não há avaliações
- GABcp 2 APROF2016 AFIMQUADRAula 1Documento8 páginasGABcp 2 APROF2016 AFIMQUADRAula 1RafaelLaurentinoAinda não há avaliações
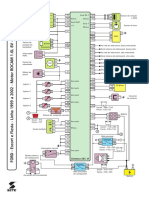
- FORD Escort Fiesta Linha 1999 A 2002 Motor ROCAM 1.6L 8V Sistema EEC VIDocumento1 páginaFORD Escort Fiesta Linha 1999 A 2002 Motor ROCAM 1.6L 8V Sistema EEC VICristianoAinda não há avaliações
- Ferramentas essenciais para eletricistasDocumento3 páginasFerramentas essenciais para eletricistasHeloisa RomãoAinda não há avaliações
- Shadowrun - Seattle SourcebookDocumento2 páginasShadowrun - Seattle SourcebookMurillo Odani de OliveiraAinda não há avaliações
- De JudIIJEF 2020 09 15 ADocumento401 páginasDe JudIIJEF 2020 09 15 AShayene RibeiroAinda não há avaliações
- Oração Do EnriquecimentoDocumento11 páginasOração Do EnriquecimentodradivinhaAinda não há avaliações
- Spink - Frezza - Práticas DiscursivasDocumento23 páginasSpink - Frezza - Práticas DiscursivasClara RodriguesAinda não há avaliações
- Gestão da Produção - Introdução e Evolução HistóricaDocumento5 páginasGestão da Produção - Introdução e Evolução Históricaanonimovw0% (1)
- Gazeta de AustinDocumento8 páginasGazeta de AustinAndre de SouzaAinda não há avaliações
- Classificação Climática B.m.aDocumento1 páginaClassificação Climática B.m.aRoberta FreireAinda não há avaliações
- IbamaDocumento17 páginasIbamaanon_700777709Ainda não há avaliações
- Aula4 2Documento37 páginasAula4 2Lucas AssisAinda não há avaliações
- Treinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNo EverandTreinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNota: 4.5 de 5 estrelas4.5/5 (169)
- Biomecânica Básica dos Exercícios: Membros InferioresNo EverandBiomecânica Básica dos Exercícios: Membros InferioresNota: 3.5 de 5 estrelas3.5/5 (8)
- Uma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNo EverandUma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNota: 4 de 5 estrelas4/5 (13)
- Atlas de anatomia: Imagens humanas e veterináriasNo EverandAtlas de anatomia: Imagens humanas e veterináriasNota: 4.5 de 5 estrelas4.5/5 (2)
- Serviços Ecossistêmicos e Planejamento Urbano: A Natureza a Favor do Desenvolvimento Sustentável das CidadesNo EverandServiços Ecossistêmicos e Planejamento Urbano: A Natureza a Favor do Desenvolvimento Sustentável das CidadesAinda não há avaliações
- Parasitologia 1: Helmintos de Interesse MédicoNo EverandParasitologia 1: Helmintos de Interesse MédicoAinda não há avaliações
- Educação ambiental: A formação do sujeito ecológicoNo EverandEducação ambiental: A formação do sujeito ecológicoAinda não há avaliações
- Equilíbrio hormonal _ Recupere equilíbrio hormonal, libido, sono e emagreça já!No EverandEquilíbrio hormonal _ Recupere equilíbrio hormonal, libido, sono e emagreça já!Nota: 5 de 5 estrelas5/5 (2)
- Fundamentos Em Cirurgia OralNo EverandFundamentos Em Cirurgia OralNota: 5 de 5 estrelas5/5 (1)
- Psicoterapia: Teorias e técnicas psicoterápicasNo EverandPsicoterapia: Teorias e técnicas psicoterápicasNota: 5 de 5 estrelas5/5 (4)
- Suplementação proteica: Aminoacidemia e saúde vascularNo EverandSuplementação proteica: Aminoacidemia e saúde vascularAinda não há avaliações
- A clínica gestáltica com adolescentes: Caminhos clínicos e institucionaisNo EverandA clínica gestáltica com adolescentes: Caminhos clínicos e institucionaisNota: 3.5 de 5 estrelas3.5/5 (3)
- Fisiologia Cardiorrespiratória Ensino SuperiorNo EverandFisiologia Cardiorrespiratória Ensino SuperiorAinda não há avaliações
- Topônimos no Sul da Bahia: nominações dos municípios originados da capitania hereditária de São Jorge dos Ilheus (1950 a 1960)No EverandTopônimos no Sul da Bahia: nominações dos municípios originados da capitania hereditária de São Jorge dos Ilheus (1950 a 1960)Nota: 5 de 5 estrelas5/5 (1)
- Concentração: Torne-se mais consciente e economize tempoNo EverandConcentração: Torne-se mais consciente e economize tempoNota: 4.5 de 5 estrelas4.5/5 (3)