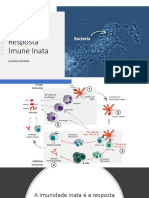
Você também pode gostar
- InflamaçãoDocumento35 páginasInflamaçãoBee NunesAinda não há avaliações
- 1250-Texto Do Artigo-2514-1-10-20121128Documento10 páginas1250-Texto Do Artigo-2514-1-10-20121128Allanis ValonAinda não há avaliações
- Células-tronco Mesenquimais Para O Tratamento Da Doença Do Enxerto Contra O Hospedeiro Aguda Resistente Aos CorticosteroidesNo EverandCélulas-tronco Mesenquimais Para O Tratamento Da Doença Do Enxerto Contra O Hospedeiro Aguda Resistente Aos CorticosteroidesAinda não há avaliações
- Aula 3. Bio 2023.1 Imunidade Inata e Inflamac807a771oDocumento79 páginasAula 3. Bio 2023.1 Imunidade Inata e Inflamac807a771oLety OliveiraAinda não há avaliações
- Artigo InflamaçãoDocumento5 páginasArtigo Inflamaçãolouisehip UFCAinda não há avaliações
- Pathophysiology of Sepsis - UpToDateDocumento27 páginasPathophysiology of Sepsis - UpToDateNetoAinda não há avaliações
- Sinalização Na Imunidade Inata e Na Inflamação: Kim Newton e Vishva M. DixitDocumento20 páginasSinalização Na Imunidade Inata e Na Inflamação: Kim Newton e Vishva M. DixitSavio SitoeAinda não há avaliações
- Pathophysiology of SepsisDocumento23 páginasPathophysiology of Sepsiscjmjyt4pbtAinda não há avaliações
- Resumo PatologiaDocumento12 páginasResumo PatologiaCelina NunesAinda não há avaliações
- Aula Inflamação - 3Documento53 páginasAula Inflamação - 3monalisa.o.maiaAinda não há avaliações
- PATOLOGIA 04 - Inflamação Crônica - MED RESUMOS - Arlindo NettoDocumento7 páginasPATOLOGIA 04 - Inflamação Crônica - MED RESUMOS - Arlindo NettoFernando Zucuni FurlanAinda não há avaliações
- Relatorio de PatologiaDocumento6 páginasRelatorio de PatologiaAndreia GomesAinda não há avaliações
- A Resolução Da Inflamação: Princípios e DesafiosDocumento1 páginaA Resolução Da Inflamação: Princípios e DesafiosTalita FernandesAinda não há avaliações
- Capítulo 5 Robbins PatologiaDocumento9 páginasCapítulo 5 Robbins PatologiaCarolina StrongerAinda não há avaliações
- 1 - Proteina C ReativaDocumento4 páginas1 - Proteina C ReativaIsabella PimentelAinda não há avaliações
- Aula 2 Hemato II Hemograma ReaciolnaisokDocumento109 páginasAula 2 Hemato II Hemograma ReaciolnaisokLudmila CaitanoAinda não há avaliações
- Aula+2+-+seção+1 2+e+1 3+Documento75 páginasAula+2+-+seção+1 2+e+1 3+Beronaldo PereiraAinda não há avaliações
- Aula11 InflamaçãoDocumento29 páginasAula11 InflamaçãoLorena Braz de OliveiraAinda não há avaliações
- FalasDocumento4 páginasFalasEnfermagem UnintaAinda não há avaliações
- Sistema Imunitário Parte IDocumento16 páginasSistema Imunitário Parte IRenata Grubert100% (1)
- KALMUNEDocumento10 páginasKALMUNELetitia GonçalvesAinda não há avaliações
- Aula 3 - Resposta Imune InataDocumento27 páginasAula 3 - Resposta Imune Inatacaroline.cabrall21Ainda não há avaliações
- Anexina1 - Famerp 13 (4) ID 144-12Documento6 páginasAnexina1 - Famerp 13 (4) ID 144-12Licinio Andrade GoncalvesAinda não há avaliações
- Proteinas de Fase AgudaDocumento5 páginasProteinas de Fase AgudaLuiz Gustavo Peron MartinsAinda não há avaliações
- Celulas Apresentadoras de Antígenos RESUMODocumento7 páginasCelulas Apresentadoras de Antígenos RESUMORosivaldo Keffer100% (1)
- Slide Resposta InflamatóriaDocumento3 páginasSlide Resposta InflamatóriaLoren MozerAinda não há avaliações
- Inflamação Aguda X CronicaDocumento2 páginasInflamação Aguda X CronicaAnderson FerreiraAinda não há avaliações
- Inflamação Crônica (Resumo)Documento3 páginasInflamação Crônica (Resumo)Hitallo BezerraAinda não há avaliações
- Resumo Imuno - Hipersensibilidade III e IV PDFDocumento7 páginasResumo Imuno - Hipersensibilidade III e IV PDFJuliana SantosAinda não há avaliações
- Inflamação e ReparoDocumento13 páginasInflamação e ReparojhonatanmichelgerhardtAinda não há avaliações
- Migracao e InflamacaoDocumento4 páginasMigracao e InflamacaoTelma FulaiAinda não há avaliações
- Inflamação AgudaDocumento38 páginasInflamação Agudakemillysouza18Ainda não há avaliações
- Problema 1 - Tutoria MadDocumento12 páginasProblema 1 - Tutoria MadVitória Amábili Laurindo RossatoAinda não há avaliações
- TEMA 2 - Anti-InflamatóriosDocumento39 páginasTEMA 2 - Anti-Inflamatóriosdiasluiza863Ainda não há avaliações
- Imunologia Dos Transplantes e TumoresDocumento24 páginasImunologia Dos Transplantes e TumoresHariel AndradeAinda não há avaliações
- Trends ImmunologyDocumento31 páginasTrends ImmunologyRogerio MachadoAinda não há avaliações
- INFLAMAÇÃODocumento18 páginasINFLAMAÇÃOfisiopaloma9Ainda não há avaliações
- Inflamação CompletaDocumento17 páginasInflamação CompletaThaís RúbioAinda não há avaliações
- Prova 1Documento5 páginasProva 1larissa rodrigues vianaAinda não há avaliações
- Sanar LESDocumento37 páginasSanar LESmanubertolAinda não há avaliações
- Inflamação - ResumoDocumento22 páginasInflamação - ResumoCynthia MunizAinda não há avaliações
- Imunologia - Imunidade InataDocumento6 páginasImunologia - Imunidade InatajessicaAinda não há avaliações
- 4 - Imunidade Inata, Inflamação, Citocinas, Migração de Leucocitos e Mecanismos Efetores CelularesDocumento11 páginas4 - Imunidade Inata, Inflamação, Citocinas, Migração de Leucocitos e Mecanismos Efetores Celularesmaria eduarda100% (2)
- A Resposta Imune Celular Da Mucosa Vaginal As VulvovaginitesDocumento3 páginasA Resposta Imune Celular Da Mucosa Vaginal As VulvovaginitesglaucodeoliveiraAinda não há avaliações
- Aula 2 Resposta InflamatóriaDocumento60 páginasAula 2 Resposta InflamatóriaDanieli Santos NascimentoAinda não há avaliações
- Resposta InflmatoriaDocumento39 páginasResposta Inflmatorialeonardo jorgeAinda não há avaliações
- 3 - AntiinflamatoriosDocumento5 páginas3 - AntiinflamatoriosGlaucia Mendes FlautoAinda não há avaliações
- ImunopatologiaDocumento9 páginasImunopatologiaOsvaldo FariaAinda não há avaliações
- Inflamação AgudaDocumento14 páginasInflamação Agudaluc100% (1)
- Aula 23 - SepseDocumento3 páginasAula 23 - SepseMarcelo GualdaAinda não há avaliações
- Livro-Texto - Unidade IIDocumento34 páginasLivro-Texto - Unidade IIRafael CedroAinda não há avaliações
- Resumao de Imunologia Clínica - Av2Documento20 páginasResumao de Imunologia Clínica - Av2leonardo SouzaAinda não há avaliações
- 5f5b8a09f60d965f3331a3b8 1633981824792Documento10 páginas5f5b8a09f60d965f3331a3b8 1633981824792Maikon FlorencianoAinda não há avaliações
- Infmação Aguda e CrônicaDocumento24 páginasInfmação Aguda e Crônicalaubaessa20Ainda não há avaliações
- Imunidade Inata e AdquiridaDocumento56 páginasImunidade Inata e AdquiridaJuan Monteiro100% (1)
- Capítulo de Síndrome Da Imunodeficiência Humana - Fernando e RodrigoDocumento73 páginasCapítulo de Síndrome Da Imunodeficiência Humana - Fernando e RodrigoJôze CataldoAinda não há avaliações
- Fasfasdffa Fasd F AfsdDocumento36 páginasFasfasdffa Fasd F AfsdfaklsdjfnasdjkfpdaskjçflkasdjAinda não há avaliações
- Sepse AtualizadaDocumento76 páginasSepse AtualizadaGiselda PachecoAinda não há avaliações
- Eucalyptus Camaldulensis É Uma: Árvore AustráliaDocumento2 páginasEucalyptus Camaldulensis É Uma: Árvore AustráliaSavio SitoeAinda não há avaliações
- Faculdade de Ciencias Direcção Académica Comprovativo de InscriçãoDocumento1 páginaFaculdade de Ciencias Direcção Académica Comprovativo de InscriçãoSavio SitoeAinda não há avaliações
- Relatório 3 BDDocumento4 páginasRelatório 3 BDSavio SitoeAinda não há avaliações
- Aula 1 Pratica BD 2023Documento8 páginasAula 1 Pratica BD 2023Savio SitoeAinda não há avaliações
- MHC SlidesDocumento40 páginasMHC Slidesalan mAinda não há avaliações
- Alergia Alimentar e QuestõesDocumento16 páginasAlergia Alimentar e QuestõesLu GaldinoAinda não há avaliações
- Aula 02.1Documento49 páginasAula 02.1Wallace SalvatoreAinda não há avaliações
- Resumo de Imunologia - Barbara Ribeiro XLVIIIDocumento16 páginasResumo de Imunologia - Barbara Ribeiro XLVIIINonato Oliveira Carvalho Junior100% (1)
- Ricardo Barbosa PDDocumento84 páginasRicardo Barbosa PDCleide FernandesAinda não há avaliações
- A Filosofia Da IntemperançaDocumento58 páginasA Filosofia Da IntemperançaWilma Matos100% (1)
- Imunologia Do Cancer PDFDocumento30 páginasImunologia Do Cancer PDFArthur CaldasAinda não há avaliações
- FEBRE HEMORRÁGICA DA DENGUE FisiopatologiaDocumento4 páginasFEBRE HEMORRÁGICA DA DENGUE FisiopatologiaLukas MadeiraAinda não há avaliações
- Unha de GatoDocumento33 páginasUnha de GatoWang HsiangAinda não há avaliações
- A Grande BatalhaDocumento15 páginasA Grande BatalhaAmin HabdidAinda não há avaliações
- BIO UNIDADE 3 (1) - JaDocumento6 páginasBIO UNIDADE 3 (1) - JajoaquinajonasAinda não há avaliações
- MEDRESUMOS 2016 - IMUNOLOGIA 10 - Ativação de Células TDocumento5 páginasMEDRESUMOS 2016 - IMUNOLOGIA 10 - Ativação de Células TneudodmAinda não há avaliações
- Resumo Aula 2 - Apresentação de AntígenoDocumento6 páginasResumo Aula 2 - Apresentação de AntígenoIngrid KasaiAinda não há avaliações
- Estudo Disciplinar Imunologia NP2Documento11 páginasEstudo Disciplinar Imunologia NP2Pri Pri100% (3)
- CELSO PEREIRA Urticaria Da Clínica À TerapeuticaDocumento394 páginasCELSO PEREIRA Urticaria Da Clínica À Terapeuticacatia_vitalAinda não há avaliações
- P1 ImunologiaDocumento21 páginasP1 Imunologializ de andradeAinda não há avaliações
- Aula Resposta PatogeniaDocumento34 páginasAula Resposta PatogeniaNinaLimaAinda não há avaliações
- Resumo Capitulo 2 AbbasDocumento5 páginasResumo Capitulo 2 AbbasDudusilAinda não há avaliações
- Gabarito Vestibular Medicina Bahiana 2019.1 2 FaseDocumento5 páginasGabarito Vestibular Medicina Bahiana 2019.1 2 FaseDermeval Vilas Boas0% (1)
- Apostila de Hematologia Biomedicina PadraoDocumento21 páginasApostila de Hematologia Biomedicina PadraoWilson RigheteAinda não há avaliações
- Cap 4 - Reconhecimento Antigênico No Sistema Imunológico AdaptativoDocumento7 páginasCap 4 - Reconhecimento Antigênico No Sistema Imunológico AdaptativoleonardoAinda não há avaliações
- Sistema Imune Do Recém-Nascido PDFDocumento8 páginasSistema Imune Do Recém-Nascido PDFLeonardo DiogoAinda não há avaliações
- Manual Crie 2006 MsDocumento158 páginasManual Crie 2006 MsFlávia MachadoAinda não há avaliações
- Lúpus Eritematoso SistêmicoDocumento36 páginasLúpus Eritematoso SistêmicoDébora Ramos de Araujo SouzaAinda não há avaliações
- Aula 13 - Imunologia Dos TransplantesDocumento56 páginasAula 13 - Imunologia Dos TransplantesvfurtadodacruzAinda não há avaliações
- Avaliando Av2 Av3Documento1 páginaAvaliando Av2 Av3Cléber FabianoAinda não há avaliações
- Aula 5 - Captura e Apresentação de Antígenos Aos Linfócitos PDFDocumento29 páginasAula 5 - Captura e Apresentação de Antígenos Aos Linfócitos PDFIris MendesAinda não há avaliações
- Imuno (2) .Docx - 20240324 - 184222 - 0000Documento30 páginasImuno (2) .Docx - 20240324 - 184222 - 0000Dhiôgo MenezesAinda não há avaliações
- Exercício de ImunologiaDocumento7 páginasExercício de ImunologiaSamuelAinda não há avaliações
- Ana Lúcia Girello - Telma Ingrid Borges de Bellis Kühn - Fundamentos Da Imuno-Hematologia Eritrocitária (2016, Senac São Paulo) - Libgen - LiDocumento172 páginasAna Lúcia Girello - Telma Ingrid Borges de Bellis Kühn - Fundamentos Da Imuno-Hematologia Eritrocitária (2016, Senac São Paulo) - Libgen - LiYuli KiaAinda não há avaliações
- Bioquímica do corpo humano: As bases moleculares do metabolismoNo EverandBioquímica do corpo humano: As bases moleculares do metabolismoAinda não há avaliações
- Treinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNo EverandTreinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNota: 4.5 de 5 estrelas4.5/5 (169)
- Heranças invisíveis do abandono afetivo: um estudo psicanalítico sobre as dimensões da experiência traumáticaNo EverandHeranças invisíveis do abandono afetivo: um estudo psicanalítico sobre as dimensões da experiência traumáticaNota: 3.5 de 5 estrelas3.5/5 (4)
- S.O.S. Autismo: Guia completo para entender o transtorno do espectro autistaNo EverandS.O.S. Autismo: Guia completo para entender o transtorno do espectro autistaNota: 4.5 de 5 estrelas4.5/5 (11)
- Os Elementos De Cura Além Dos MedicamentosNo EverandOs Elementos De Cura Além Dos MedicamentosNota: 4.5 de 5 estrelas4.5/5 (3)
- RISCOS, VULNERABILIDADES E CONDICIONANTES URBANOSNo EverandRISCOS, VULNERABILIDADES E CONDICIONANTES URBANOSAinda não há avaliações
- Psicoterapia: Teorias e técnicas psicoterápicasNo EverandPsicoterapia: Teorias e técnicas psicoterápicasNota: 5 de 5 estrelas5/5 (4)
- Remineralização de Solos com Rochas Ricas em Silício, Potássio e Outros Nutrientes: Uma Solução para Solos Pouco Férteis e Envelhecidos de Cerrados?No EverandRemineralização de Solos com Rochas Ricas em Silício, Potássio e Outros Nutrientes: Uma Solução para Solos Pouco Férteis e Envelhecidos de Cerrados?Ainda não há avaliações
- Uma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNo EverandUma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNota: 4 de 5 estrelas4/5 (13)
- Aprender mais rápido: Habilidades de estudo, memória e neurociênciaNo EverandAprender mais rápido: Habilidades de estudo, memória e neurociênciaNota: 4 de 5 estrelas4/5 (5)
- Transtorno de Personalidade Borderline Mais de 30 segredos para retomar sua vida Ao lidar com TPBNo EverandTranstorno de Personalidade Borderline Mais de 30 segredos para retomar sua vida Ao lidar com TPBNota: 2.5 de 5 estrelas2.5/5 (3)
- Dieta Cetogênica: Seu Guia Completo E Definitivo Para Perder Peso Para IniciantesNo EverandDieta Cetogênica: Seu Guia Completo E Definitivo Para Perder Peso Para IniciantesNota: 5 de 5 estrelas5/5 (1)
- Biomecânica Básica dos Exercícios: Membros InferioresNo EverandBiomecânica Básica dos Exercícios: Membros InferioresNota: 3.5 de 5 estrelas3.5/5 (8)