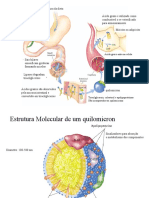
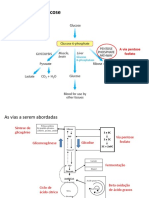
Você também pode gostar
- Metabolismo LIPIDEOSDocumento20 páginasMetabolismo LIPIDEOSVanderval SIlva de OliveiraAinda não há avaliações
- Glicolise, Gliconeogenese E A Via Das Pentoses-Fosfato - Parte 2 PDFDocumento5 páginasGlicolise, Gliconeogenese E A Via Das Pentoses-Fosfato - Parte 2 PDFAna YooAinda não há avaliações
- Ebook - Descomplicando A Bioquimica PDFDocumento101 páginasEbook - Descomplicando A Bioquimica PDFEdmara Silva100% (2)
- AD2 Bioquímica IIDocumento2 páginasAD2 Bioquímica IIGabriel MagalhãesAinda não há avaliações
- 1740718 (2).pptDocumento20 páginas1740718 (2).pptAle Araujo100% (1)
- Biossíntese de LipídiosDocumento7 páginasBiossíntese de LipídiosLeonardo Peixoto Carvalho100% (1)
- Aula 2 - Fisiopatologia e Diagnóstico Laboratorial Das Dislipidemias 2016.1Documento22 páginasAula 2 - Fisiopatologia e Diagnóstico Laboratorial Das Dislipidemias 2016.1Jailton MagalhãesAinda não há avaliações
- Ficha Trabalho FotossínteseDocumento2 páginasFicha Trabalho FotossíntesePaula Lopes100% (1)
- Resumo Bioquímica Ciclo Da UreiaDocumento4 páginasResumo Bioquímica Ciclo Da UreiamatheusAinda não há avaliações
- BIOENERGÉTICADocumento45 páginasBIOENERGÉTICAAdriana100% (4)
- Resumo de Ciclo de KrebsDocumento6 páginasResumo de Ciclo de KrebsLívia Lopes100% (2)
- 6 - Oxidação de Aminoácidos e Ciclo Da Ureia ResumoDocumento7 páginas6 - Oxidação de Aminoácidos e Ciclo Da Ureia ResumoCaruma GuerreiroAinda não há avaliações
- Transaminação de AADocumento7 páginasTransaminação de AABeatriz Souza LimaAinda não há avaliações
- Metabolismo de AminoácidosDocumento4 páginasMetabolismo de AminoácidosKarina MoreiraAinda não há avaliações
- Metabolismo de AminoácidosDocumento2 páginasMetabolismo de AminoácidosJúliaAinda não há avaliações
- Pontos Importantes No Metabolismo de ProteínasDocumento10 páginasPontos Importantes No Metabolismo de ProteínasMariana SilvaAinda não há avaliações
- Metabolismo Do NitrogênioDocumento14 páginasMetabolismo Do NitrogênioVandik CandidoAinda não há avaliações
- Nitrogen I o Raquel MDocumento13 páginasNitrogen I o Raquel MgeizeAinda não há avaliações
- Ciclo Da UreiaDocumento11 páginasCiclo Da UreiaThais Gama100% (1)
- RevisãoDocumento6 páginasRevisãoe236610Ainda não há avaliações
- Além de Serem Constituintes Das Proteínas Os Aminoácidos Podem Ser Usados Como Precursores de Moléculas Biológicas AzotadasDocumento5 páginasAlém de Serem Constituintes Das Proteínas Os Aminoácidos Podem Ser Usados Como Precursores de Moléculas Biológicas AzotadasJunea DrewsAinda não há avaliações
- Metabolismo de ProteínasDocumento26 páginasMetabolismo de ProteínasGuilherme ZulimAinda não há avaliações
- Ciclo Da Ureia - ResumoDocumento4 páginasCiclo Da Ureia - ResumoArlindo FernandesAinda não há avaliações
- Aula 7 - Degradação Dos Alfa-CetoácidosDocumento5 páginasAula 7 - Degradação Dos Alfa-CetoácidosArielle MoraesAinda não há avaliações
- Bioquímica: Aula 4Documento16 páginasBioquímica: Aula 4Tuani de MirandaAinda não há avaliações
- Aula 6 - Metabolismo Dos AminoácidosDocumento6 páginasAula 6 - Metabolismo Dos AminoácidosArielle MoraesAinda não há avaliações
- 6-Ciclo Da UreiaDocumento14 páginas6-Ciclo Da Ureiaapi-27441481Ainda não há avaliações
- Metabolismo Dos AminoácidosDocumento7 páginasMetabolismo Dos AminoácidosErica AdrianoAinda não há avaliações
- Degradação de AminoácidosDocumento4 páginasDegradação de AminoácidosSabrina XavierAinda não há avaliações
- A AlaninaDocumento5 páginasA AlaninaSara Krebold100% (1)
- Ciclo Da UréiaDocumento20 páginasCiclo Da UréiaPriscila Kelly Domingos100% (1)
- Resumos BCMDocumento12 páginasResumos BCMRaquel MotaAinda não há avaliações
- Sessão 7Documento5 páginasSessão 7Gabriel PiresAinda não há avaliações
- Metabolismo de ProteinasDocumento2 páginasMetabolismo de ProteinasMirelleTavaresAinda não há avaliações
- (Resumo) Bioquim - Catabolismo de ProteinasDocumento16 páginas(Resumo) Bioquim - Catabolismo de Proteinasabresser100% (2)
- Ciclo Da UreiaDocumento65 páginasCiclo Da Ureiamellogenilza88Ainda não há avaliações
- Estou A Partilhar o Ficheiro Exame 2016 Bioquimica Perguntas Consigo - 210129 - 011144Documento5 páginasEstou A Partilhar o Ficheiro Exame 2016 Bioquimica Perguntas Consigo - 210129 - 01114428485Ainda não há avaliações
- AMINOÁCIDOSDocumento19 páginasAMINOÁCIDOSRodolfo CrespoAinda não há avaliações
- Degradação de Triagliceróis e Ácidos GraxosDocumento11 páginasDegradação de Triagliceróis e Ácidos GraxosJosceane PereiraAinda não há avaliações
- G11 - Catabolismo Do Azoto Dos AminoácidosDocumento13 páginasG11 - Catabolismo Do Azoto Dos Aminoácidosup202007257Ainda não há avaliações
- Metabolismo de AminoácidosDocumento8 páginasMetabolismo de AminoácidosDenisar PalmitoAinda não há avaliações
- Ciclo Da UreiaDocumento9 páginasCiclo Da UreiaJandderson CostaAinda não há avaliações
- A Oxidação e A Biossíntese Dos Ácidos GraxosDocumento2 páginasA Oxidação e A Biossíntese Dos Ácidos GraxosCarlos MarianiAinda não há avaliações
- Ciclo Do Ácido CítricoDocumento14 páginasCiclo Do Ácido CítricoEry AlencarAinda não há avaliações
- Ciclo Da UreiaDocumento6 páginasCiclo Da UreiaSabrina Nicole100% (1)
- P4 - ArmazenamentoDocumento14 páginasP4 - ArmazenamentoPedro Henrique Almeida de BritoAinda não há avaliações
- Oxidação de Aminoácidos e Produção de UreiaDocumento10 páginasOxidação de Aminoácidos e Produção de UreiaErika BennaiaAinda não há avaliações
- Atividade de BioquímicaDocumento4 páginasAtividade de BioquímicaRenata BarrosoAinda não há avaliações
- Estudo Dirigido (Síntese de AG, Hormônios... )Documento2 páginasEstudo Dirigido (Síntese de AG, Hormônios... )Mariana Gonçalves MacedoAinda não há avaliações
- Lista de AminoácidosDocumento2 páginasLista de Aminoácidos86t2fgkbtnAinda não há avaliações
- Estudo Dirigido de Ciclo Da Uréia - BioquímicaDocumento3 páginasEstudo Dirigido de Ciclo Da Uréia - BioquímicaLucas FeitosaAinda não há avaliações
- Roteiro Elisa Aula 6 - Metabolismo de Aminoácidos e Ciclo Da UréiaDocumento8 páginasRoteiro Elisa Aula 6 - Metabolismo de Aminoácidos e Ciclo Da UréiaElisa de AlmeidaAinda não há avaliações
- Degradação e Biossíntese de AminoácidosDocumento34 páginasDegradação e Biossíntese de AminoácidosAndréa MouraAinda não há avaliações
- Apostila Gliclise Ciclo Do Cido Ctrico Cadeia Transportadora de EletronsDocumento13 páginasApostila Gliclise Ciclo Do Cido Ctrico Cadeia Transportadora de Eletronsemailinutil2manAinda não há avaliações
- Resumos de BioquimicaDocumento42 páginasResumos de BioquimicaAna Carolina Soares EstevesAinda não há avaliações
- Metabolismo Dos Aminoácidos e Ciclo Do Citrato. ResumoDocumento1 páginaMetabolismo Dos Aminoácidos e Ciclo Do Citrato. ResumoLuana Maciel PaivaAinda não há avaliações
- Aula 4 - Ciclo Da UreiaDocumento22 páginasAula 4 - Ciclo Da UreiaBruno Moreira Dos SantosAinda não há avaliações
- Os Aminoácidos Não São Armazenados Pelo CorpoDocumento6 páginasOs Aminoácidos Não São Armazenados Pelo CorpoadrianokotAinda não há avaliações
- Aula 10 - Metabolismo de Proteinas (Aula Remota)Documento11 páginasAula 10 - Metabolismo de Proteinas (Aula Remota)Marcos PauloAinda não há avaliações
- Metabolismo Dos Aminoácidos MonografiaDocumento3 páginasMetabolismo Dos Aminoácidos MonografiaDianaSantosAinda não há avaliações
- Ciclo KrebsDocumento25 páginasCiclo KrebsLuís VeigaAinda não há avaliações
- Metabolismo de LipídiosDocumento3 páginasMetabolismo de LipídiosPatrícia BentherAinda não há avaliações
- 2º Teste de Bioenergética e Metabolismo Correcção 2Documento5 páginas2º Teste de Bioenergética e Metabolismo Correcção 2Carlos CostaAinda não há avaliações
- Ciclo Da Ureia Ou Ciclo Da Ornitina, GUIOMAR, IsABELDocumento4 páginasCiclo Da Ureia Ou Ciclo Da Ornitina, GUIOMAR, IsABELadilsonjacinto19945Ainda não há avaliações
- Catabolismo de AaDocumento48 páginasCatabolismo de AaJoão MariaAinda não há avaliações
- Aula 2 Bioquímica Da Nutrição e Atividade FísicaDocumento60 páginasAula 2 Bioquímica Da Nutrição e Atividade FísicaMileny AugustoAinda não há avaliações
- Exercício Respiração e FermentaçãoDocumento2 páginasExercício Respiração e FermentaçãoGabriel FagundesAinda não há avaliações
- 5 - Sistemas de Fornecimento de Energia - ATPCP PDFDocumento3 páginas5 - Sistemas de Fornecimento de Energia - ATPCP PDFLuiz Eduardo RodriguesAinda não há avaliações
- Aula 3.8 Via Pentose FosfatoDocumento20 páginasAula 3.8 Via Pentose Fosfatowalas joãoAinda não há avaliações
- Sônia LopesDocumento10 páginasSônia LopesRomario SousaAinda não há avaliações
- Ciclo de KrebsDocumento2 páginasCiclo de KrebsRenato SouzaAinda não há avaliações
- Aula 18 - Ciclos de Krebs e GlioxilatoDocumento62 páginasAula 18 - Ciclos de Krebs e GlioxilatoWeza RicardoAinda não há avaliações
- Processos Biológicos: Capítulo 3 - de Onde Vem A Energia para As Células Funcionarem?Documento45 páginasProcessos Biológicos: Capítulo 3 - de Onde Vem A Energia para As Células Funcionarem?Alan OliveiraAinda não há avaliações
- 5-Sistema Anaeróbio LáticoDocumento18 páginas5-Sistema Anaeróbio LáticoRaphaella RodriguesAinda não há avaliações
- Aula 10 - o Ciclo Do Ácido Cítrico p.2Documento24 páginasAula 10 - o Ciclo Do Ácido Cítrico p.2juliana moraes niloAinda não há avaliações
- Via Do GlucuronatoDocumento4 páginasVia Do Glucuronatoapi-27441481Ainda não há avaliações
- Slides Aula 11. CTE e Fosforilação OxidativaDocumento26 páginasSlides Aula 11. CTE e Fosforilação OxidativaÂngelo AbnerAinda não há avaliações
- 1.estudo Dirigido Glicolise e Via Das PentosesDocumento1 página1.estudo Dirigido Glicolise e Via Das PentosesMonieli VialiAinda não há avaliações
- Fermentação e Respiração AeróbiaDocumento5 páginasFermentação e Respiração AeróbiaCatarina CarmoAinda não há avaliações
- Ionilda ResultadoDocumento1 páginaIonilda ResultadoFabiano QuadrosAinda não há avaliações
- 6 - Metabolismo de Carboidratos I - Glicólise e GliconeogêneseDocumento8 páginas6 - Metabolismo de Carboidratos I - Glicólise e GliconeogêneseFelipe RostirollaAinda não há avaliações
- Quiz MetabolismoDocumento3 páginasQuiz Metabolismopholl.gregorioAinda não há avaliações
- Prof. José Guilherme Prado Martin Aula 03: Metabolismo Microbiano I: Obtenção de EnergiaDocumento43 páginasProf. José Guilherme Prado Martin Aula 03: Metabolismo Microbiano I: Obtenção de Energiadanilo CanettieriAinda não há avaliações
- Mitocôndria Conversão Energética e Respiração CelularDocumento23 páginasMitocôndria Conversão Energética e Respiração CelularLiliane LevoratoAinda não há avaliações
- AD1 Bioquímica 2021-2Documento2 páginasAD1 Bioquímica 2021-2Lue Wyn FirAinda não há avaliações
- Aula 5 - MitocôndriasDocumento20 páginasAula 5 - MitocôndriasvictoriaapolinarioAinda não há avaliações
- Semienem-Biologia1-Metabolismo Energético-15-06-2020Documento9 páginasSemienem-Biologia1-Metabolismo Energético-15-06-2020Ana Luíza RibeiroAinda não há avaliações
- Biologia 15-06-2012 Metabolismo EnergéticoDocumento15 páginasBiologia 15-06-2012 Metabolismo EnergéticoLucas MendesAinda não há avaliações