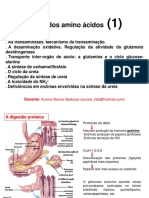
Você também pode gostar
- Degradacao Dos Acidos Graxos 2020Documento60 páginasDegradacao Dos Acidos Graxos 2020Bio FontesAinda não há avaliações
- Lipólise e B-OxidaçãoDocumento26 páginasLipólise e B-OxidaçãoFernando FernandesAinda não há avaliações
- B-Oxidaçao de LipídeosDocumento45 páginasB-Oxidaçao de LipídeosbolotatiagoAinda não há avaliações
- Metabolismo LIPIDEOSDocumento20 páginasMetabolismo LIPIDEOSVanderval SIlva de OliveiraAinda não há avaliações
- B OxidacaoDocumento49 páginasB OxidacaoMariana Gonçalves MacedoAinda não há avaliações
- Aula Revisao Metabolismo CArboidratosDocumento32 páginasAula Revisao Metabolismo CArboidratosmonekobeAinda não há avaliações
- Aula Revisao Metabolismo CArboidratosDocumento32 páginasAula Revisao Metabolismo CArboidratosmonekobeAinda não há avaliações
- Introdução Ao Metabolismo - Metabolismo de Carboidrato - UBCDocumento101 páginasIntrodução Ao Metabolismo - Metabolismo de Carboidrato - UBCLeiliane RibeiroAinda não há avaliações
- Bioquímica - Aula 12 - Metabolismo Dos LipídiosDocumento29 páginasBioquímica - Aula 12 - Metabolismo Dos LipídiosTássio BrunoAinda não há avaliações
- BioqDocumento30 páginasBioqMichelle BarcellosAinda não há avaliações
- 002GLICOLISEDocumento21 páginas002GLICOLISEAna Carla Peixoto100% (1)
- Revisão p2 de BoinhaDocumento45 páginasRevisão p2 de BoinhaGlenda FrançaAinda não há avaliações
- BIOENERGÉTICA - CópiaDocumento37 páginasBIOENERGÉTICA - CópiaMaria Luíza AguiarAinda não há avaliações
- Aula 5 - Oxidacao Acidos Graxos 2021Documento24 páginasAula 5 - Oxidacao Acidos Graxos 2021Maria Carolina Azêdo MurussiAinda não há avaliações
- AULA 3 - Metabolismo de CHO No ExercicioDocumento48 páginasAULA 3 - Metabolismo de CHO No ExercicioTaise TavaresAinda não há avaliações
- Módulo 6 - Oxidação de Ácidos GraxosDocumento17 páginasMódulo 6 - Oxidação de Ácidos GraxosBeatriz De Sousa CoelhoAinda não há avaliações
- Ciclo de KrebsDocumento2 páginasCiclo de KrebsAlana FreitasAinda não há avaliações
- Ciclo de KrebsDocumento42 páginasCiclo de KrebsDaniela BrumAinda não há avaliações
- Ciclo de Krebs ENGDocumento25 páginasCiclo de Krebs ENGGabriela Ruwer AmaralAinda não há avaliações
- 05 - Resumo RespiracaoDocumento2 páginas05 - Resumo RespiracaoLara MartinsAinda não há avaliações
- Superesumodebiokill 3 BDocumento3 páginasSuperesumodebiokill 3 BJohanaAinda não há avaliações
- Aula 8-9 - Respiração Celular e FermentaçãoDocumento77 páginasAula 8-9 - Respiração Celular e FermentaçãoWesleyJúniorAinda não há avaliações
- Beta Oxidaoremoto2020Documento25 páginasBeta Oxidaoremoto2020amanda abreuAinda não há avaliações
- Apostila de BioquímicaDocumento13 páginasApostila de BioquímicaLívia ChristineAinda não há avaliações
- Aula Introd A MetabolismoDocumento95 páginasAula Introd A Metabolismoluci cristinaAinda não há avaliações
- Aula II Metabolismo CarboidratosDocumento63 páginasAula II Metabolismo CarboidratosBila BernardesAinda não há avaliações
- Metabolismo Degradativo e Bioquímico Dos AçúcaresDocumento17 páginasMetabolismo Degradativo e Bioquímico Dos AçúcaresLucas P.Ainda não há avaliações
- Lista de Exercicios Aula 13-Ciclo de Krebs e GlicogênioDocumento7 páginasLista de Exercicios Aula 13-Ciclo de Krebs e GlicogênioMaria MoreiraAinda não há avaliações
- Aula Sistemas EnergeticosDocumento79 páginasAula Sistemas EnergeticosAndré Guelli Trainer IAinda não há avaliações
- BIOQUIMICADocumento7 páginasBIOQUIMICAWellerson SardiAinda não há avaliações
- 2.respiração CelularDocumento24 páginas2.respiração CelularCristiana GuedesAinda não há avaliações
- Resumo Biologia Celular e MolecularDocumento21 páginasResumo Biologia Celular e Molecularcarolina cardosoAinda não há avaliações
- Aula Biossíntese de Lipídeos - Enf - 2023Documento31 páginasAula Biossíntese de Lipídeos - Enf - 2023Arthur NascimentoAinda não há avaliações
- 1668558229694+metabolismo Dos LipídiosDocumento2 páginas1668558229694+metabolismo Dos LipídiosAnnieChase 2018Ainda não há avaliações
- Estudo Dirigido 4 Met2017 RespostasDocumento2 páginasEstudo Dirigido 4 Met2017 RespostasThailana Silva SousaAinda não há avaliações
- CDP3Documento7 páginasCDP3gab.francis2Ainda não há avaliações
- Fundamentos Da Bioquímica - BioenergeticaDocumento54 páginasFundamentos Da Bioquímica - BioenergeticamonekobeAinda não há avaliações
- Bioquímica - Introdução Ao Metabolismo e EnzimasDocumento29 páginasBioquímica - Introdução Ao Metabolismo e EnzimasMariana LinsAinda não há avaliações
- Aila - 3 - Metabolismo de Lipídeos PDFDocumento9 páginasAila - 3 - Metabolismo de Lipídeos PDFVinícius DearmasAinda não há avaliações
- Beta Oxidação 2017Documento44 páginasBeta Oxidação 2017Maria Eduarda Frota100% (1)
- Beta OxidaçãoDocumento43 páginasBeta OxidaçãoRique EmanuelAinda não há avaliações
- Metabolismo - Energetico RespiraçãoDocumento46 páginasMetabolismo - Energetico RespiraçãowerottydesluizAinda não há avaliações
- 2º Teste de Bioenergética e Metabolismo Correcção 2Documento5 páginas2º Teste de Bioenergética e Metabolismo Correcção 2Carlos CostaAinda não há avaliações
- Aula 9 - Glicólise e Destinos Do PiruvatoDocumento53 páginasAula 9 - Glicólise e Destinos Do PiruvatoFernando BritoAinda não há avaliações
- Aula - Metabolismo de LipídioDocumento17 páginasAula - Metabolismo de Lipídiojuliana moraes niloAinda não há avaliações
- Respiração CelularDocumento14 páginasRespiração CelularMANOEL PEREIRA DA SILVA JUNIORAinda não há avaliações
- Gabarito de Bioquímica - Aula 5Documento3 páginasGabarito de Bioquímica - Aula 5Michelle BarcellosAinda não há avaliações
- Semana 12 - RespiraçãoDocumento19 páginasSemana 12 - RespiraçãoElizabethCristinaAinda não há avaliações
- Bioquimica C.K.Documento8 páginasBioquimica C.K.EronAinda não há avaliações
- Nutrição e Metabolismo Ciclo Do Ácido CítricoDocumento25 páginasNutrição e Metabolismo Ciclo Do Ácido CítricoAlexia Braz BomfimAinda não há avaliações
- Aula 10 - Metabolismo LipídeosDocumento24 páginasAula 10 - Metabolismo LipídeosValeskaSena100% (1)
- MetabolismoDocumento55 páginasMetabolismoRichard WooliteAinda não há avaliações
- Ciclo de KrebsESA2023.1Documento23 páginasCiclo de KrebsESA2023.1lukinhasfranco790Ainda não há avaliações
- LipoliseDocumento5 páginasLipoliseeabonacc100% (2)
- Bioquimica MetabólicaDocumento83 páginasBioquimica MetabólicaEdgard Freitas100% (1)
- Processos Bioenergéticos e NeoglicogêneseDocumento50 páginasProcessos Bioenergéticos e NeoglicogêneseyvignoloAinda não há avaliações
- Respiração AeróbiaDocumento19 páginasRespiração AeróbiaScherzbergAinda não há avaliações
- AT17 CicloKrebsDocumento30 páginasAT17 CicloKrebsMateus GarciaAinda não há avaliações
- Glicólise X GliconeogêneseDocumento14 páginasGlicólise X GliconeogênesejaraAinda não há avaliações
- ComprovativoDocumento2 páginasComprovativoAfonso Henriques NunesAinda não há avaliações
- Corticoterapia SistémicaDocumento5 páginasCorticoterapia SistémicaAfonso Henriques NunesAinda não há avaliações
- Abordagem de Adenopatias e Esplenomegalia - NotasPraticasDocumento21 páginasAbordagem de Adenopatias e Esplenomegalia - NotasPraticasAfonso Henriques NunesAinda não há avaliações
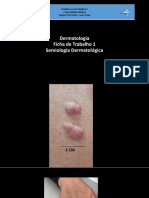
- Derma FichaTrabalho1Documento27 páginasDerma FichaTrabalho1Afonso Henriques NunesAinda não há avaliações
- Aula 12Documento19 páginasAula 12Afonso Henriques NunesAinda não há avaliações
- Cancro Da Mama Cirurgia II VerónicaDocumento29 páginasCancro Da Mama Cirurgia II VerónicaAfonso Henriques NunesAinda não há avaliações
- Caso Gastro 2Documento8 páginasCaso Gastro 2Afonso Henriques NunesAinda não há avaliações
- Aula 5Documento9 páginasAula 5Afonso Henriques NunesAinda não há avaliações
- AS1 Aula 4 28set18 TemporalDocumento54 páginasAS1 Aula 4 28set18 TemporalAfonso Henriques NunesAinda não há avaliações
- Manual - Injecao - Injepro s3000Documento52 páginasManual - Injecao - Injepro s3000Alex RodriguesAinda não há avaliações
- Bomba Pressurizada RowaDocumento20 páginasBomba Pressurizada RowafernandowfrancaAinda não há avaliações
- Caracterizacao Da Nafta Petroquimica para Producao de Produtos AromaticosDocumento141 páginasCaracterizacao Da Nafta Petroquimica para Producao de Produtos Aromaticoszeqs9Ainda não há avaliações
- Eclipse Lunar e SolarDocumento15 páginasEclipse Lunar e Solarvictor hugo brAinda não há avaliações
- Relatório Aço CK45 e ST37.2Documento32 páginasRelatório Aço CK45 e ST37.2Mónica Pinheiro0% (1)
- Material Aulão MaceióDocumento10 páginasMaterial Aulão MaceióEdnardoQuintilianoCabralFilhoAinda não há avaliações
- Mestre Mário Sassi - No Limiar Do Terceiro Milênio 01Documento81 páginasMestre Mário Sassi - No Limiar Do Terceiro Milênio 01Jaciara Celina100% (1)
- Fisica-1ano ETEASDDocumento4 páginasFisica-1ano ETEASDValter DantasAinda não há avaliações
- Lista de Exercícios 2Documento2 páginasLista de Exercícios 22023313743Ainda não há avaliações
- Computação Gráfica 2Documento49 páginasComputação Gráfica 2Karine SantosAinda não há avaliações
- Naturalismo e Existencialismo Na Filosofia de HareDocumento8 páginasNaturalismo e Existencialismo Na Filosofia de HarecrisejoelAinda não há avaliações
- Uniforme Sesi Modelo 2022Documento60 páginasUniforme Sesi Modelo 2022Bia CcassitaAinda não há avaliações
- Protocolo DHCPDocumento2 páginasProtocolo DHCPifilomenaAinda não há avaliações
- Descritivo Nomencaltura de Programa o StarSoftDocumento13 páginasDescritivo Nomencaltura de Programa o StarSoftGilson RamosAinda não há avaliações
- Bomba Multitec 6 EstagiosDocumento39 páginasBomba Multitec 6 EstagiosPaulo VieiraAinda não há avaliações
- A Importância Da Lógica e Da Argumentação para Os Profissionais Do DireitoDocumento3 páginasA Importância Da Lógica e Da Argumentação para Os Profissionais Do DireitoIvi GalvãoAinda não há avaliações
- Exercícios de Cinemática 01Documento4 páginasExercícios de Cinemática 01Marcos GuilhermeAinda não há avaliações
- Como Calcular o CPVDocumento12 páginasComo Calcular o CPVreginaldo.contabil7730Ainda não há avaliações
- Toc - tcm2 - Total Cost Manegement - TRADUÇÃO PTDocumento71 páginasToc - tcm2 - Total Cost Manegement - TRADUÇÃO PTLucindo SouzaAinda não há avaliações
- Seminário Interdisciplinar 3Documento3 páginasSeminário Interdisciplinar 3Temistocles Dias de Oliveira NetoAinda não há avaliações
- EEAR 3º SimuladoDocumento4 páginasEEAR 3º SimuladoAldo MouraAinda não há avaliações
- Manual NdriveDocumento39 páginasManual NdriveJoão Paulo VieiraAinda não há avaliações
- Citröen Peugeot - 1.8 16VDocumento6 páginasCitröen Peugeot - 1.8 16VMauro VasconcelosAinda não há avaliações
- Mecânica Da Fratura EscritaDocumento1 páginaMecânica Da Fratura EscritaMisael SoutoAinda não há avaliações
- Aula-04 ENE005 PDFDocumento32 páginasAula-04 ENE005 PDFVirgilio RosaAinda não há avaliações
- Ficha Teorica 12a - Modulo de Um Numero Real. S.gabrielDocumento16 páginasFicha Teorica 12a - Modulo de Um Numero Real. S.gabriellourenapaulaAinda não há avaliações
- FMC BR 20180802222103 PDFDocumento492 páginasFMC BR 20180802222103 PDFfelipeAinda não há avaliações
- Aula 9 - Prof. Jeferson FraytagDocumento16 páginasAula 9 - Prof. Jeferson Fraytageramos_77Ainda não há avaliações
- Ed 05Documento14 páginasEd 05PAPAI SMURFAinda não há avaliações
- Mudanças de Estado Físico Da MatériaDocumento1 páginaMudanças de Estado Físico Da MatériaSolange Alves de MatosAinda não há avaliações