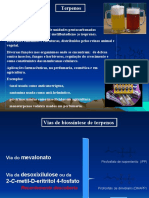
Você também pode gostar
- Os principais tipos de terpenoides classificados pelo número de unidades de isoprenoDocumento13 páginasOs principais tipos de terpenoides classificados pelo número de unidades de isoprenoMatheus AlmeidaAinda não há avaliações
- Farmag 2Documento95 páginasFarmag 2zilmar100% (2)
- Terpenóides: compostos derivados de isoprenoDocumento26 páginasTerpenóides: compostos derivados de isoprenoCelso da Silva100% (1)
- Sebenta Gnosia2Documento70 páginasSebenta Gnosia2nandaff5100% (2)
- Terpenos - Métodos de Extração, Sua Formação e AplicaçõesDocumento70 páginasTerpenos - Métodos de Extração, Sua Formação e AplicaçõescweltonfelipeAinda não há avaliações
- Hidrocarbonetos em Ponte, Espiranos e TerpénicosDocumento19 páginasHidrocarbonetos em Ponte, Espiranos e Terpénicosapi-27441481100% (3)
- 1C Lípidos TerpenosDocumento25 páginas1C Lípidos Terpenosapi-27441481100% (2)
- TrabalhoDocumento21 páginasTrabalhoNatalino Manuel RapiequeAinda não há avaliações
- 7 - Terpenos PDFDocumento60 páginas7 - Terpenos PDFJota CarlosAinda não há avaliações
- As Biomoléculas São Moléculas Resultantes Dos MetabolismosDocumento2 páginasAs Biomoléculas São Moléculas Resultantes Dos MetabolismosJoão GabrielAinda não há avaliações
- Classes de Substancias Presentes Nos Oleo EssenciaisDocumento6 páginasClasses de Substancias Presentes Nos Oleo EssenciaismjmorcelliAinda não há avaliações
- Bicos pulverizadores: tipos e usos em aplicações agrícolasDocumento24 páginasBicos pulverizadores: tipos e usos em aplicações agrícolasdiegoAinda não há avaliações
- TerpenosDocumento11 páginasTerpenosJota CarlosAinda não há avaliações
- Aula 02 - Terpenoides e EsteroidesDocumento29 páginasAula 02 - Terpenoides e EsteroidesRoberta Cordeiro100% (1)
- 9705-Texto Do Artigo-36865-1-10-20200309Documento14 páginas9705-Texto Do Artigo-36865-1-10-20200309Ingredy Brunele100% (1)
- 2b-Lignanas, Ligninas e Suberização: Tecido Suberizado Tecido LenhificadoDocumento26 páginas2b-Lignanas, Ligninas e Suberização: Tecido Suberizado Tecido LenhificadoMatheus AlmeidaAinda não há avaliações
- PDF-BM2-aula 9TDocumento31 páginasPDF-BM2-aula 9TMatheus AlmeidaAinda não há avaliações
- Farmacognosia Óleos EssenciaisDocumento20 páginasFarmacognosia Óleos EssenciaisvitrinetrenddAinda não há avaliações
- Óleos essenciais, terpenos e esteróides na Farmacognosia PuraDocumento50 páginasÓleos essenciais, terpenos e esteróides na Farmacognosia PuraRichardson AlbuquerqueAinda não há avaliações
- Farmacognosia - Aula 03 - Metabolismo VegetalDocumento39 páginasFarmacognosia - Aula 03 - Metabolismo VegetalFabio ViniciosAinda não há avaliações
- Óleos essenciais: propriedades, obtenção e usosDocumento60 páginasÓleos essenciais: propriedades, obtenção e usosLarissa LopesAinda não há avaliações
- Diterpenos e Triterpenos - João e Rogério - Versão Final 2Documento18 páginasDiterpenos e Triterpenos - João e Rogério - Versão Final 2João GabrielAinda não há avaliações
- QUIMICA DOS PRODUTOS NATURAISDocumento20 páginasQUIMICA DOS PRODUTOS NATURAISAndreia FilipaAinda não há avaliações
- BIOQUIMICA - Terpenos e TerpenóidesDocumento35 páginasBIOQUIMICA - Terpenos e TerpenóidesClarice Guizoni50% (4)
- Aula 06 -Transformação de Energia na CélulaDocumento53 páginasAula 06 -Transformação de Energia na Célulajulianafurtado2103Ainda não há avaliações
- Monoterpenos e MonoterpenóidesDocumento57 páginasMonoterpenos e MonoterpenóidesVinícius FernandesAinda não há avaliações
- TerpenosDocumento5 páginasTerpenosThays Ramos100% (1)
- FotossínteseDocumento6 páginasFotossíntesebeatriz.barbosa.0607Ainda não há avaliações
- Prova IDocumento18 páginasProva IuandersonkAinda não há avaliações
- Aula MetabolismoDocumento12 páginasAula MetabolismoPatricia SilvaAinda não há avaliações
- Lista de Exercícios - Nomenclatura de HidrocarbonetosDocumento2 páginasLista de Exercícios - Nomenclatura de HidrocarbonetosalvescolonamariaeduardaAinda não há avaliações
- Metabolismo Dos NucleotídeosDocumento3 páginasMetabolismo Dos NucleotídeosBreno S. GonçalvesAinda não há avaliações
- Oleos Essenciais Essencias-1Documento33 páginasOleos Essenciais Essencias-1Diego ChicanelAinda não há avaliações
- Óleos essenciais: propriedades e métodos de extraçãoDocumento33 páginasÓleos essenciais: propriedades e métodos de extraçãoDiego ChicanelAinda não há avaliações
- Genetic ADocumento12 páginasGenetic AkewinwilliansAinda não há avaliações
- Plastídios e cloroplastos: estrutura e funçãoDocumento24 páginasPlastídios e cloroplastos: estrutura e funçãohuigorAinda não há avaliações
- Extração do Óleo Essencial do Limão CravoDocumento13 páginasExtração do Óleo Essencial do Limão CravoCarlos EduardoAinda não há avaliações
- Ver Nos Meus SlidesDocumento13 páginasVer Nos Meus SlidesJoao CostaAinda não há avaliações
- Trabalho Polímeros OficialDocumento29 páginasTrabalho Polímeros OficialRennan MarinhoAinda não há avaliações
- Polímeros e BiomoléculasDocumento28 páginasPolímeros e BiomoléculasGiseleCristina40% (5)
- Fotossíntes (Parte 1)Documento23 páginasFotossíntes (Parte 1)Renato GonçalvesAinda não há avaliações
- Introdução Ao Metabolismo de Lipídeos CompletaDocumento54 páginasIntrodução Ao Metabolismo de Lipídeos Completaotacide RosarioAinda não há avaliações
- Questo Es - Fotossi NteseDocumento16 páginasQuesto Es - Fotossi NteseHeloísa CristinaAinda não há avaliações
- Terpenos e esteroides: classificação, biossíntese e propriedadesDocumento30 páginasTerpenos e esteroides: classificação, biossíntese e propriedadesJoab Ana Menezes100% (1)
- Fotossíntese ImprimirDocumento4 páginasFotossíntese ImprimirRosineide ParenteAinda não há avaliações
- Res01-Replicação Do DNADocumento3 páginasRes01-Replicação Do DNAhumbertosouza1alvesAinda não há avaliações
- Metabólitos Secundários Via Ácido ChiquímicoDocumento10 páginasMetabólitos Secundários Via Ácido ChiquímicoLuis Eduardo DornelesAinda não há avaliações
- Introdução aos Polímeros: Propriedades, Tipos e PolimerizaçãoDocumento56 páginasIntrodução aos Polímeros: Propriedades, Tipos e PolimerizaçãoDiego Arturo PintoAinda não há avaliações
- Macromoléculas celularesDocumento60 páginasMacromoléculas celularesMarcella KarolineAinda não há avaliações
- Os microrganismos metanogênicos são os que produzem metano como parte integral de seu metabolismo energético. Eles são anaeróbios obrigatórios. A maioria são mesofíticos, ou seja, a temperatura ótima de crescimento éDocumento1 páginaOs microrganismos metanogênicos são os que produzem metano como parte integral de seu metabolismo energético. Eles são anaeróbios obrigatórios. A maioria são mesofíticos, ou seja, a temperatura ótima de crescimento éGabriely LemeAinda não há avaliações
- Hormônios da Tireóide: Função e BiossínteseDocumento5 páginasHormônios da Tireóide: Função e BiossínteseCarla FunatoAinda não há avaliações
- Polímeros e BiomoléculasDocumento38 páginasPolímeros e Biomoléculaslc788095Ainda não há avaliações
- DNA e síntese proteicaDocumento38 páginasDNA e síntese proteicaSoraia MartinsAinda não há avaliações
- Aula 2 - AlcalóidesDocumento10 páginasAula 2 - AlcalóidesGiovanna Saadeh NevesAinda não há avaliações
- Processamento do RNAmDocumento44 páginasProcessamento do RNAmWesley Luzetti FotoranAinda não há avaliações
- Resumo Do Livro Medical Herbalism HoffmannDocumento2 páginasResumo Do Livro Medical Herbalism Hoffmannbiomed.becaAinda não há avaliações
- Genética Médica para Médicos e Estudantes de Medicina: OncologiaNo EverandGenética Médica para Médicos e Estudantes de Medicina: OncologiaAinda não há avaliações
- Flavonoides ProtocoloDocumento2 páginasFlavonoides ProtocoloMatheus AlmeidaAinda não há avaliações
- Plantas medicinais: da pré-história à era modernaDocumento28 páginasPlantas medicinais: da pré-história à era modernaMatheus AlmeidaAinda não há avaliações
- PDF-BM2-aula 9TDocumento31 páginasPDF-BM2-aula 9TMatheus AlmeidaAinda não há avaliações
- PDF-BM2-aula 8TDocumento18 páginasPDF-BM2-aula 8TMatheus AlmeidaAinda não há avaliações
- Bioquímica Metabolismo IiDocumento7 páginasBioquímica Metabolismo IiMatheus AlmeidaAinda não há avaliações
- 2b-Lignanas, Ligninas e Suberização: Tecido Suberizado Tecido LenhificadoDocumento26 páginas2b-Lignanas, Ligninas e Suberização: Tecido Suberizado Tecido LenhificadoMatheus AlmeidaAinda não há avaliações
- 2c-Cumarinas, Estilbe-Nos, Estirilpironas, e ArilpironasDocumento10 páginas2c-Cumarinas, Estilbe-Nos, Estirilpironas, e ArilpironasMatheus AlmeidaAinda não há avaliações
- Método filosófico vs científicoDocumento1 páginaMétodo filosófico vs científicoMatheus AlmeidaAinda não há avaliações
- Metabolitos SecundáriosDocumento34 páginasMetabolitos SecundáriosMatheus AlmeidaAinda não há avaliações
- Distonia Focal Do Músico e A Nova Craniopuntura de YamamotoDocumento19 páginasDistonia Focal Do Músico e A Nova Craniopuntura de YamamotoLivia CarolinaAinda não há avaliações
- Introdução Psicologia: Definição e Evolução EstudoDocumento14 páginasIntrodução Psicologia: Definição e Evolução EstudoJoão José AraújoAinda não há avaliações
- Bula MetiêDocumento2 páginasBula MetiêJULIANA MACEDOAinda não há avaliações
- Cópia de Cardapio 2023 Pizzaria Rocha - 20231003 - 234111 - 0000-1Documento4 páginasCópia de Cardapio 2023 Pizzaria Rocha - 20231003 - 234111 - 0000-1Marcelo Landi BassoAinda não há avaliações
- Uso Da Matriz PIER No Estudo Dos Problemas Ecológicos UrbanosDocumento10 páginasUso Da Matriz PIER No Estudo Dos Problemas Ecológicos UrbanosSergio Alfredo MacoreAinda não há avaliações
- Mitose e Meiose (Aula Extra)Documento3 páginasMitose e Meiose (Aula Extra)Aline Aparecida Bastos PortelaAinda não há avaliações
- Tintas naturais: elaboração e aplicação em oficinas na UFPRDocumento11 páginasTintas naturais: elaboração e aplicação em oficinas na UFPRJosé Contigio JuniorAinda não há avaliações
- Platao. Hipias MaiorDocumento53 páginasPlatao. Hipias MaiorNico AraujoAinda não há avaliações
- Moisés e a jornada de Israel pelo desertoDocumento44 páginasMoisés e a jornada de Israel pelo desertoCarolina MarinhoAinda não há avaliações
- Atividade 1.1 - Exercício de FixaçãoDocumento4 páginasAtividade 1.1 - Exercício de FixaçãoLorráynny GuimarãesAinda não há avaliações
- Ficha de EV. BiológicaDocumento2 páginasFicha de EV. Biológicacat famousAinda não há avaliações
- Anatomia Humana IDocumento76 páginasAnatomia Humana IAllanDeivityDosSantos100% (4)
- Cadeias, Pirâmides e Teias Alimentares - BiologiaDocumento1 páginaCadeias, Pirâmides e Teias Alimentares - BiologiaThaís LoiolaAinda não há avaliações
- Modulo Economia AmbientalDocumento87 páginasModulo Economia AmbientalSérgio SaguarAinda não há avaliações
- Fundações profundas: tipos de estacasDocumento16 páginasFundações profundas: tipos de estacasd23m89Ainda não há avaliações
- Concurso Público para Professor de Educação BásicaDocumento12 páginasConcurso Público para Professor de Educação Básicaduda_tricolorAinda não há avaliações
- Critérios AdjudicaçãoDocumento31 páginasCritérios AdjudicaçãoLuís AlexandreAinda não há avaliações
- A Caixa de Ferramentas de DeusDocumento4 páginasA Caixa de Ferramentas de Deuswalter_pacheco_1Ainda não há avaliações
- Manual Do Sensor de PresençaDocumento1 páginaManual Do Sensor de PresençaGloria ZeladaAinda não há avaliações
- Pinagem Ultimate ECU Test Microplex 613ADocumento4 páginasPinagem Ultimate ECU Test Microplex 613AJapao MillerAinda não há avaliações
- Aposentadoria por invalidez - laudo pericialDocumento13 páginasAposentadoria por invalidez - laudo pericialcarlos henrique nunes pintoAinda não há avaliações
- Conhecimentos Técnicos de Helicópteros (Carateristicas Gerais)Documento58 páginasConhecimentos Técnicos de Helicópteros (Carateristicas Gerais)Raizen Urameshi100% (1)
- Eti Catálogo EtiledDocumento44 páginasEti Catálogo EtiledVEMATELAinda não há avaliações
- Manual de Normas e Rotinas Da Unidade Materno Infantil (UMI) - Clínica ObstétricaDocumento25 páginasManual de Normas e Rotinas Da Unidade Materno Infantil (UMI) - Clínica ObstétricaLauraTechy100% (1)
- Introducao A Epistemologia Da CienciaDocumento172 páginasIntroducao A Epistemologia Da CienciaRogerio Andrade de Avila0% (1)
- Consciencia CulinariaDocumento3 páginasConsciencia CulinariaAnderson Kleber50% (2)
- O. G. de Carvalho: vida e obra do escritor piauienseDocumento24 páginasO. G. de Carvalho: vida e obra do escritor piauienseedilsonsfonteneleAinda não há avaliações
- MAN-002 - Manutenção de Máquinas e Equipamentos de Produção Rev 12 19-03-2013Documento6 páginasMAN-002 - Manutenção de Máquinas e Equipamentos de Produção Rev 12 19-03-2013Jefferson MoraesAinda não há avaliações
- Dom Juan - Jessica LuizDocumento227 páginasDom Juan - Jessica LuizMj MenegattiAinda não há avaliações
- Patologia Veterinária Geral: Causas, Sinais e TratamentosDocumento15 páginasPatologia Veterinária Geral: Causas, Sinais e TratamentosMilenaAinda não há avaliações