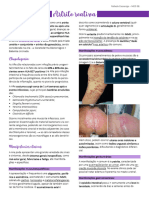
Você também pode gostar
- Autoanticorpos após uso de terapia biológica: estudo multicêntrico brasileiroNo EverandAutoanticorpos após uso de terapia biológica: estudo multicêntrico brasileiroAinda não há avaliações
- AR - Celulas B - 230427 - 105931Documento14 páginasAR - Celulas B - 230427 - 105931Larissa HayneAinda não há avaliações
- Proteínas de Fase AgudaDocumento31 páginasProteínas de Fase AgudaTiago VidigalAinda não há avaliações
- 14.1 - Osteomielite AgudaDocumento7 páginas14.1 - Osteomielite AgudacarvalhalgiuliaAinda não há avaliações
- ARDocumento4 páginasARedvelcaAinda não há avaliações
- AR - Celulas T - 230427 - 110052Documento10 páginasAR - Celulas T - 230427 - 110052Larissa HayneAinda não há avaliações
- ReumatologiaDocumento52 páginasReumatologiaInês Gonçalves FreitasAinda não há avaliações
- Aterosclerose, Inflamação e Infecção: Artigo de RevisãoDocumento7 páginasAterosclerose, Inflamação e Infecção: Artigo de Revisãofernandaneves30Ainda não há avaliações
- Patogênese Da Artrite ReumatoideDocumento29 páginasPatogênese Da Artrite ReumatoideLeonardo Tavares de Oliveira100% (1)
- Artrite Reactiva Por Giardia Lambia Num Doente Com Défict de IgE Secretora PDFDocumento5 páginasArtrite Reactiva Por Giardia Lambia Num Doente Com Défict de IgE Secretora PDFEnzoAinda não há avaliações
- Artrite ReumatoideDocumento11 páginasArtrite ReumatoideRafaela FreitasAinda não há avaliações
- Diagnóstico e Tratamento de Choque Séptico em CãesDocumento5 páginasDiagnóstico e Tratamento de Choque Séptico em CãesAlberto De Oliveira BrandãoAinda não há avaliações
- Atrite ReumatoideDocumento5 páginasAtrite Reumatoideeduardobadin92Ainda não há avaliações
- Artrite Reativa - RafaDocumento3 páginasArtrite Reativa - RafaMarco Aurélio Joslin AugustoAinda não há avaliações
- Páginas de Rockwood Fraturas em Adultos, 8ed7Documento51 páginasPáginas de Rockwood Fraturas em Adultos, 8ed7Estevão CananAinda não há avaliações
- Síndrome Da Resposta Inflamatória Sistêmica - SepseDocumento7 páginasSíndrome Da Resposta Inflamatória Sistêmica - SepseAndre TatiAinda não há avaliações
- Marcadores InflamatoriosDocumento34 páginasMarcadores InflamatoriosAurilane Oliveira da SilvaAinda não há avaliações
- Uveite e EADocumento5 páginasUveite e EAJayce Layana CallouAinda não há avaliações
- Artrite ReativaDocumento16 páginasArtrite ReativasfstucaAinda não há avaliações
- J Ocl 2016 12 008 en PTDocumento8 páginasJ Ocl 2016 12 008 en PTLeonardo BarrosoAinda não há avaliações
- Livro Fundamentos de Imunologia Do Roitt Cap Online ExtraDocumento43 páginasLivro Fundamentos de Imunologia Do Roitt Cap Online ExtraEder OliveiraAinda não há avaliações
- Artrite ReumatoideDocumento114 páginasArtrite Reumatoidecarolina cassianoAinda não há avaliações
- Imunossenescencia 1Documento8 páginasImunossenescencia 1ArieleAinda não há avaliações
- Problema 3 - Modulo 1 - Artrite ReumatóideDocumento18 páginasProblema 3 - Modulo 1 - Artrite ReumatóideLuiz BelarminoAinda não há avaliações
- Aula Prática 1 PDFDocumento10 páginasAula Prática 1 PDFManoell Netto100% (1)
- Sanar LESDocumento37 páginasSanar LESmanubertolAinda não há avaliações
- Características e Diagnóstico Diferencial Das Doenças Inflamatórias IntestinaisDocumento7 páginasCaracterísticas e Diagnóstico Diferencial Das Doenças Inflamatórias IntestinaisIlfran IIAinda não há avaliações
- Wallach 10ed Interpretação de Exames LaboratoriaisDocumento7 páginasWallach 10ed Interpretação de Exames Laboratoriaislarissa0% (1)
- 22702c-DC - Manif Cutaneas Princip DC ReumaticasDocumento26 páginas22702c-DC - Manif Cutaneas Princip DC ReumaticasBruno CavalcanteAinda não há avaliações
- Reumatologia - Questões ComentadasDocumento23 páginasReumatologia - Questões ComentadasCarol MedtoAinda não há avaliações
- Medcel - Reumatologia: LesDocumento16 páginasMedcel - Reumatologia: LesPerola NakAinda não há avaliações
- Unifip - ImunologiaDocumento22 páginasUnifip - ImunologiamylenaalvesnobregaAinda não há avaliações
- Roteiro-de-Estudo ARDocumento6 páginasRoteiro-de-Estudo ARRafaela FreitasAinda não há avaliações
- Patologias ReumatológicasDocumento93 páginasPatologias ReumatológicasÉrica OliveiraAinda não há avaliações
- Dçs AutoimunespptxDocumento27 páginasDçs AutoimunespptxTainá CostaAinda não há avaliações
- 775 1643 1 SMDocumento8 páginas775 1643 1 SMLucas PereiraAinda não há avaliações
- DIP - MeningitesDocumento5 páginasDIP - MeningitesEdson FernandesAinda não há avaliações
- GabaritoDocumento22 páginasGabaritoTais LopesAinda não há avaliações
- LúpusDocumento8 páginasLúpusIsabela BanderaliAinda não há avaliações
- Imunologia e EnvelhecimentoDocumento8 páginasImunologia e EnvelhecimentomarjoriemelyssaAinda não há avaliações
- Pioderma GangrenosoDocumento4 páginasPioderma GangrenosoDennysson CorreiaAinda não há avaliações
- Arginina PDFDocumento7 páginasArginina PDFMaiara BotossoAinda não há avaliações
- Relatorio Final LohannaDocumento31 páginasRelatorio Final LohannaLohanna Menezes BatistaAinda não há avaliações
- Aula 2.2 - Análise Crítica Dos Exames COMPLETODocumento23 páginasAula 2.2 - Análise Crítica Dos Exames COMPLETOlucasmateriais2024Ainda não há avaliações
- Exames de Sangue para ReumatismoDocumento33 páginasExames de Sangue para ReumatismoRicardo Fernandes de PaulaAinda não há avaliações
- Artrite ReumatoideDocumento10 páginasArtrite ReumatoideManassés AlmeidaAinda não há avaliações
- A Importância Do Profissional Farmacêutico No Aconselhamento Ao Portador de Lupus Eritematoso PDFDocumento7 páginasA Importância Do Profissional Farmacêutico No Aconselhamento Ao Portador de Lupus Eritematoso PDFSílvia HelenaAinda não há avaliações
- Artigo Esclerose SistemicaDocumento3 páginasArtigo Esclerose Sistemicacamila ziganti favaroAinda não há avaliações
- Aftose Revisão BibliográficaDocumento8 páginasAftose Revisão BibliográficaEduardo AzevedoAinda não há avaliações
- Atividade Integradora - P9M2: 1 - Compreender A Fisiopatologia Da Sepse e Choque SépticoDocumento11 páginasAtividade Integradora - P9M2: 1 - Compreender A Fisiopatologia Da Sepse e Choque SépticoGeorge LucasAinda não há avaliações
- Entesite de Calcaneo e EspondilopatiasDocumento4 páginasEntesite de Calcaneo e EspondilopatiasCelia AlcantaraAinda não há avaliações
- AR SanarflixDocumento22 páginasAR SanarflixPedro XavierAinda não há avaliações
- Doencas ReumatologicasDocumento66 páginasDoencas ReumatologicasLaercio Dezinho100% (1)
- Aula - Artrite ReumatoideDocumento33 páginasAula - Artrite ReumatoideEdyllaSilvaAinda não há avaliações
- 2 PBDocumento5 páginas2 PBLaila SiqueiraAinda não há avaliações
- CASO 10 - Vulvovaginites e Artrite ReumatóideDocumento9 páginasCASO 10 - Vulvovaginites e Artrite Reumatóidegelyson.diasAinda não há avaliações
- Aim 1 Reumato T10Documento31 páginasAim 1 Reumato T10fevmateusAinda não há avaliações
- Artrite Reumatoide (AR) Um Simples GuiaDocumento7 páginasArtrite Reumatoide (AR) Um Simples GuiaL1PE8Ainda não há avaliações
- Linfoma e Espondilite Anquilosante Ea em Uso de AnDocumento1 páginaLinfoma e Espondilite Anquilosante Ea em Uso de AnVictor Dias AlmeidaAinda não há avaliações
- Encefalomielite AviáriaDocumento1 páginaEncefalomielite AviáriaKaline DiasAinda não há avaliações
- Agente - Administrativo 03-01-23Documento13 páginasAgente - Administrativo 03-01-23Gabriel SaraivaAinda não há avaliações
- Planner JJ 5Documento3 páginasPlanner JJ 5Geraldo FerreiraAinda não há avaliações
- Apostila Saude MentalDocumento356 páginasApostila Saude MentalProf. Bira Ilustrador100% (1)
- Riscos Na Marcenaria - Izabella DinizDocumento2 páginasRiscos Na Marcenaria - Izabella DinizIzabella DinizAinda não há avaliações
- Jeny USDocumento8 páginasJeny USGiullia PeronAinda não há avaliações
- A Proteção Constitucional Das Pessoas Portadoras de Deficiência - Luiz Alberto David AraujoDocumento90 páginasA Proteção Constitucional Das Pessoas Portadoras de Deficiência - Luiz Alberto David AraujoGiovanna MDPAinda não há avaliações
- NR 07 - Saúde Mental e Trabalho 09Documento20 páginasNR 07 - Saúde Mental e Trabalho 09CPSSTAinda não há avaliações
- Aula V - NivelamentoDocumento2 páginasAula V - NivelamentoLuiz Ferreira JuniorAinda não há avaliações
- Provas Anteriores Eags Sef Sem Gabarito-1Documento221 páginasProvas Anteriores Eags Sef Sem Gabarito-1Renata SilvaAinda não há avaliações
- Semana 3Documento2 páginasSemana 3Vitorino CoelhoAinda não há avaliações
- LipomaDocumento6 páginasLipomaCaio GonçalvesAinda não há avaliações
- Modelo Pre Projeto Completo Enfermagem V0Documento11 páginasModelo Pre Projeto Completo Enfermagem V0Tatiane Carvalho Confessor ConfessorAinda não há avaliações
- GabaritoDocumento32 páginasGabaritoAmaury VieiraAinda não há avaliações
- ARTIGO - Corpo e Automultilação - Um Estudo de CasoDocumento9 páginasARTIGO - Corpo e Automultilação - Um Estudo de CasoMariana Luz PatezAinda não há avaliações
- Interpretacao de Texto Cafe Uma Bebida Natural e Saudavel 2º Ano Do Ensino Medio Modelo EditavelDocumento2 páginasInterpretacao de Texto Cafe Uma Bebida Natural e Saudavel 2º Ano Do Ensino Medio Modelo EditavelMadalena dos Santos BarbosaAinda não há avaliações
- POP 12 - 2016 - Fisioterapia - Cardiovascular - Ambulatorial - Corrigido Versão FinalDocumento21 páginasPOP 12 - 2016 - Fisioterapia - Cardiovascular - Ambulatorial - Corrigido Versão Finallis costaAinda não há avaliações
- Procedimento Anestésico:: Capítulo 7Documento26 páginasProcedimento Anestésico:: Capítulo 7Jose Mattos100% (1)
- Meios de CulturaDocumento12 páginasMeios de CulturaPaulo HenriqueAinda não há avaliações
- PDF 20211214 111309 0000Documento96 páginasPDF 20211214 111309 0000Thainara Lina Alves dos Reis100% (1)
- Marlon Correa de Oliveira: 52574449 José Ricardo Brizzi Chiani DR ( ) : CRMDocumento6 páginasMarlon Correa de Oliveira: 52574449 José Ricardo Brizzi Chiani DR ( ) : CRMMarlon correa de olveiraAinda não há avaliações
- Radiologia em EndodontiaDocumento2 páginasRadiologia em EndodontiaMichelle Souza100% (1)
- Caderneta Da Saude - 2017 - Menina PDFDocumento96 páginasCaderneta Da Saude - 2017 - Menina PDFSantos MrnAinda não há avaliações
- Alcolismo em IdosoDocumento17 páginasAlcolismo em IdosoFábio VinholyAinda não há avaliações
- Termo de Consentimento Livre e EsclarecidoDocumento2 páginasTermo de Consentimento Livre e EsclarecidoAna Paula Cunha67% (3)
- Guia Genética Humana Unidade 3Documento13 páginasGuia Genética Humana Unidade 3Jeová MadeiroAinda não há avaliações
- Proteína C Reativa - Ultrassensível: SinônimosDocumento2 páginasProteína C Reativa - Ultrassensível: SinônimosSthefane S FerreiraAinda não há avaliações
- Slides Taping+Estetico INAESPDocumento133 páginasSlides Taping+Estetico INAESPFlavia Daniela100% (2)
- Medicina Veterinária Preventiva - Aula 1Documento19 páginasMedicina Veterinária Preventiva - Aula 1Rodolfo mktAinda não há avaliações
- Dificuldades de AprendizagemDocumento13 páginasDificuldades de AprendizagemChauale Da Linda ChameAinda não há avaliações
- O Grande Livro Da LinfaDocumento31 páginasO Grande Livro Da LinfaDayane Costa 2Ainda não há avaliações