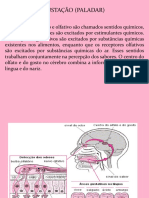
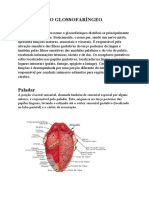
Você também pode gostar
- Aula 15 - Estesiologia - O Império Dos SentidosDocumento14 páginasAula 15 - Estesiologia - O Império Dos SentidosThais Mello100% (5)
- Olfato e PaladarDocumento16 páginasOlfato e PaladarAmanda NathalyAinda não há avaliações
- Capítulo 53 - Os Sentidos Químicos - Gustação e Olfação - 3 PáginasDocumento3 páginasCapítulo 53 - Os Sentidos Químicos - Gustação e Olfação - 3 PáginasandreamokadaAinda não há avaliações
- Aula Anatomia e Fisiologia Da Gustação e OlfaçãoDocumento28 páginasAula Anatomia e Fisiologia Da Gustação e OlfaçãoLiliane PanAinda não há avaliações
- Sistema Sensorial Nos AnimaisDocumento57 páginasSistema Sensorial Nos AnimaisNhofa Delgado100% (5)
- Atenção e Cuidado Da Saúde Bucal Da Pessoa Com DeficiênciaDocumento232 páginasAtenção e Cuidado Da Saúde Bucal Da Pessoa Com DeficiênciaMayara França100% (1)
- Aula Paladar Olfato e TatoDocumento33 páginasAula Paladar Olfato e TatoOdilon Calian50% (2)
- Sentido Gustativo e OlfativoDocumento6 páginasSentido Gustativo e OlfativoCaroline SantanaAinda não há avaliações
- 09 Sentido Gustativo OlfativoDocumento6 páginas09 Sentido Gustativo OlfativoDaniel Delani100% (1)
- Trabalho Sobre o OlfatoDocumento10 páginasTrabalho Sobre o OlfatoLucas Barbosa SoaresAinda não há avaliações
- Receptores Gustativos e OlfatoriosDocumento4 páginasReceptores Gustativos e OlfatoriosTainar VieiraAinda não há avaliações
- 5 - Sentidos Quimicos, Olfação e GustaçãoDocumento6 páginas5 - Sentidos Quimicos, Olfação e GustaçãoRafael LopesAinda não há avaliações
- 2 TicsDocumento5 páginas2 TicsBeatriz GoulartAinda não há avaliações
- Fisiologia - 05 - Órgãos Dos Sentidos - Gustação e OlfaçãoDocumento31 páginasFisiologia - 05 - Órgãos Dos Sentidos - Gustação e OlfaçãoMatheusAinda não há avaliações
- Sistema Estomatognatico ArtigoDocumento4 páginasSistema Estomatognatico ArtigoFúlvia NicollyAinda não há avaliações
- Trabalho Paladar ProntoDocumento17 páginasTrabalho Paladar Prontonaycg50% (2)
- Diferença de ReceptoresDocumento3 páginasDiferença de ReceptoresIsaac AlencarAinda não há avaliações
- Orgaos Dos SentidosDocumento35 páginasOrgaos Dos SentidosJuliana Ventura100% (1)
- O OLFATO EscritoDocumento4 páginasO OLFATO EscritoJose RibeiroAinda não há avaliações
- Órgãos Dos SentidosDocumento41 páginasÓrgãos Dos SentidosPatricia SousaAinda não há avaliações
- SentidosDocumento8 páginasSentidosIsabelle CeruttiAinda não há avaliações
- Sistema SensorialDocumento6 páginasSistema SensorialFernando CarvalhoAinda não há avaliações
- Sistema SensorialDocumento26 páginasSistema SensorialDario Cipriano Silverio100% (1)
- Diferença de Receptores IsaacDocumento3 páginasDiferença de Receptores IsaacIsaac AlencarAinda não há avaliações
- Aula 04 - Fisio I 2023Documento85 páginasAula 04 - Fisio I 2023Krain SanMeloAinda não há avaliações
- Região OlfatóriaDocumento2 páginasRegião OlfatóriaJoão Paulo VergeAinda não há avaliações
- Resumo Da Lingua Sabor e PapilasDocumento2 páginasResumo Da Lingua Sabor e PapilasMarilay ZacAinda não há avaliações
- Sensações QuímicasDocumento20 páginasSensações QuímicasmicalAinda não há avaliações
- Sistema Olfativo ComponentesDocumento6 páginasSistema Olfativo ComponentesviniciusAinda não há avaliações
- Sistema Sensorial NovoDocumento61 páginasSistema Sensorial NovoAnonymous 85j0VSCvAinda não há avaliações
- Gustação e Olfação - EC2019Documento53 páginasGustação e Olfação - EC2019Lucas GustavoAinda não há avaliações
- 3 - Gosto e OlfatoDocumento6 páginas3 - Gosto e OlfatobeatrizAinda não há avaliações
- Ciencias 6oano C Maa 24072020.docxDocumento3 páginasCiencias 6oano C Maa 24072020.docxEdna LopesAinda não há avaliações
- Trabalho Eduardo-1Documento14 páginasTrabalho Eduardo-1Eduardo AraoAinda não há avaliações
- OlfatoDocumento20 páginasOlfatoJailaneCostaAinda não há avaliações
- Fisiologia Da Gustação e OlfatoDocumento2 páginasFisiologia Da Gustação e OlfatoJulia GoicoecheaAinda não há avaliações
- ExperimentoDocumento1 páginaExperimentoArthur SampaioAinda não há avaliações
- A GustaçãoDocumento6 páginasA GustaçãoPriscila MendesAinda não há avaliações
- Orgãos SensoriaisDocumento2 páginasOrgãos SensoriaisDanielle MariaAinda não há avaliações
- Quimiorrecepção - Luiza e EduardoDocumento47 páginasQuimiorrecepção - Luiza e EduardoRoberto SantosAinda não há avaliações
- Olfa CaoDocumento27 páginasOlfa CaoOsório TrigoAinda não há avaliações
- Órganos de Los SentidosDocumento79 páginasÓrganos de Los SentidosZu MendozaAinda não há avaliações
- Aula Anatomia Sensorial EnfermagemDocumento95 páginasAula Anatomia Sensorial EnfermagemPedro Augusto100% (1)
- O PaladarDocumento16 páginasO PaladarMuvane CanzerAinda não há avaliações
- Sistema Sensorial O.G.T.A 2019 6 PDFDocumento25 páginasSistema Sensorial O.G.T.A 2019 6 PDFSimone MaiaAinda não há avaliações
- Processos Psicológicos Básicos (PPB) - Olfato + PaladarDocumento16 páginasProcessos Psicológicos Básicos (PPB) - Olfato + Paladarsephiroth1208Ainda não há avaliações
- Os Orgãos Do Sentido e Seus Mecanismo de FuncionamentoDocumento39 páginasOs Orgãos Do Sentido e Seus Mecanismo de FuncionamentoTaizi RodriguesAinda não há avaliações
- Fisiologia Da GustaçãoDocumento12 páginasFisiologia Da GustaçãoGabriel MartinsAinda não há avaliações
- Fisiologia AnimalDocumento5 páginasFisiologia AnimalCamila MoraisAinda não há avaliações
- Glossofaríngeo Funções CorrigidoDocumento4 páginasGlossofaríngeo Funções CorrigidoLívia DuarteAinda não há avaliações
- QuimiorreceptoresDocumento10 páginasQuimiorreceptoresTainá PestanaAinda não há avaliações
- Problema 04, Sistema SensorialDocumento11 páginasProblema 04, Sistema SensorialIgor HenriqueAinda não há avaliações
- Sistema SensorialDocumento24 páginasSistema SensorialAna Paula BatistaAinda não há avaliações
- RESUMODocumento8 páginasRESUMOEstudos VitóriaAinda não há avaliações
- Formação Anatômica Dos Órgãos e Sentidos Do Sistema SensorialDocumento7 páginasFormação Anatômica Dos Órgãos e Sentidos Do Sistema Sensorialjoyce cristina pereiraAinda não há avaliações
- Os Cinco Sentidos Do Corpo HumanoDocumento9 páginasOs Cinco Sentidos Do Corpo HumanoNoemí MaiaAinda não há avaliações
- Faculdade Anhanguera Somestesico 2Documento13 páginasFaculdade Anhanguera Somestesico 2Jéssica LaportaAinda não há avaliações
- GustaçãoDocumento2 páginasGustaçãoANNA BEATRIZ MINGA ZANARDIAinda não há avaliações
- Como parar de Roncar de modo natural!: Dormir melhor e se livrar do ronco agora!No EverandComo parar de Roncar de modo natural!: Dormir melhor e se livrar do ronco agora!Ainda não há avaliações
- Apg 7 - Sistema Autonomo e LimbicoDocumento13 páginasApg 7 - Sistema Autonomo e LimbicoAnita DamkeAinda não há avaliações
- Tick S 4Documento1 páginaTick S 4Anita DamkeAinda não há avaliações
- Tick S 2Documento1 páginaTick S 2Anita DamkeAinda não há avaliações
- Apg 6 - Vias Eferentes, Cereblo e Nucleo de BaseDocumento21 páginasApg 6 - Vias Eferentes, Cereblo e Nucleo de BaseAnita DamkeAinda não há avaliações
- Tick S 3Documento1 páginaTick S 3Anita DamkeAinda não há avaliações
- Tick S 1Documento1 páginaTick S 1Anita DamkeAinda não há avaliações
- Tick S 3Documento1 páginaTick S 3Anita DamkeAinda não há avaliações
- APG 3 - Coluna, Medula Espinal e Arco-ReflexoDocumento24 páginasAPG 3 - Coluna, Medula Espinal e Arco-ReflexoAnita DamkeAinda não há avaliações
- Resumos+Apgs+ +Soi+II+Afya+ +parte+1Documento199 páginasResumos+Apgs+ +Soi+II+Afya+ +parte+1Anita DamkeAinda não há avaliações
- Resumos+Apgs+ +Soi+II+Afya+ +parte+1Documento199 páginasResumos+Apgs+ +Soi+II+Afya+ +parte+1Anita DamkeAinda não há avaliações
- Gabarito Exercícios Sistema SensorialDocumento5 páginasGabarito Exercícios Sistema SensorialikazueAinda não há avaliações
- Testes AcumétricosDocumento35 páginasTestes AcumétricosRenata BarrosAinda não há avaliações
- Paga o Trabalh o Dr.Documento38 páginasPaga o Trabalh o Dr.lucas mayungulaAinda não há avaliações
- Morfologia Externa PDFDocumento37 páginasMorfologia Externa PDFDiego CamposAinda não há avaliações
- Slide Aparelho AuditivoDocumento9 páginasSlide Aparelho AuditivoJéssica SantanaAinda não há avaliações
- Audicao 3Documento301 páginasAudicao 3Alice FerminoAinda não há avaliações
- Orgaos Dos SentidosDocumento35 páginasOrgaos Dos SentidosJuliana Ventura100% (1)
- Integração de Segurança No Trabalho - NR-18Documento78 páginasIntegração de Segurança No Trabalho - NR-18cunha cunhaAinda não há avaliações
- Livro Educação Especial Deficiencia Auditiva Vol.iDocumento303 páginasLivro Educação Especial Deficiencia Auditiva Vol.iRenata ModestoAinda não há avaliações
- TONTURASDocumento10 páginasTONTURASsofiarcolomboAinda não há avaliações
- Seminario Sistema VestibularDocumento34 páginasSeminario Sistema Vestibularkarinapdias2612Ainda não há avaliações
- O Ruído Nas Obras Da Construção Civil e o Risco de Surdez OcupacionalDocumento148 páginasO Ruído Nas Obras Da Construção Civil e o Risco de Surdez OcupacionalAlexsandra PennaAinda não há avaliações
- Fdocumentos - Tips Orgaos Do Sentido Audicao Tato Paladar Visao Olfato Sistema SensorialDocumento118 páginasFdocumentos - Tips Orgaos Do Sentido Audicao Tato Paladar Visao Olfato Sistema SensorialEdileide JesusAinda não há avaliações
- Curso Básico de PSICOACUSTICADocumento15 páginasCurso Básico de PSICOACUSTICAzybegy8Ainda não há avaliações
- Manual Tas TatDocumento122 páginasManual Tas TatromeonederlandsAinda não há avaliações
- Resumo Palestra Vestibular Ronaldo Casimiro 2019Documento3 páginasResumo Palestra Vestibular Ronaldo Casimiro 2019Julia Maria dos SantosAinda não há avaliações
- QUESTÕES DE HISTOLOGIA DOS ORGAOS DOS SENTIDOS - Trabalho Do Victor FaculdadeDocumento3 páginasQUESTÕES DE HISTOLOGIA DOS ORGAOS DOS SENTIDOS - Trabalho Do Victor FaculdadeUederson Aragão da SilvaAinda não há avaliações
- Apostila Sonoplastia LucasDocumento44 páginasApostila Sonoplastia LucasBruno Alvim100% (1)
- Anatomia Da OrelhaDocumento11 páginasAnatomia Da OrelhaRafael Amadeu Barbosa Luperi100% (1)
- Questionario Sistema AuditivoDocumento2 páginasQuestionario Sistema Auditivobeatriz0mslsAinda não há avaliações
- CORTEXIDocumento22 páginasCORTEXISérgio WahhabAinda não há avaliações
- 2013.11.08 - Bases Físicas Da AudiçãoDocumento39 páginas2013.11.08 - Bases Físicas Da Audiçãomaizinha0810Ainda não há avaliações
- Fisiologia Básica Da AudiçãoDocumento5 páginasFisiologia Básica Da AudiçãoCaio AmaralAinda não há avaliações
- Termos Técnicos Da Fonoaudiologia ..Documento8 páginasTermos Técnicos Da Fonoaudiologia ..bs696132Ainda não há avaliações
- Livro - Eixo 2 - Cirurgiões-Dentistas PDFDocumento232 páginasLivro - Eixo 2 - Cirurgiões-Dentistas PDFVitoria100% (2)
- Libras PDFDocumento73 páginasLibras PDFHerica Ramos100% (1)
- Sistema Biométrico para Reconhecimento de Orador Pela Segmentação Convex HullDocumento53 páginasSistema Biométrico para Reconhecimento de Orador Pela Segmentação Convex HulldamibardAinda não há avaliações
- Física Acústica para FonoaudiologiaDocumento8 páginasFísica Acústica para FonoaudiologiafilipecwaAinda não há avaliações
- Portifólio AnatomofisiologiaDocumento41 páginasPortifólio AnatomofisiologiaFernando MonteiroAinda não há avaliações