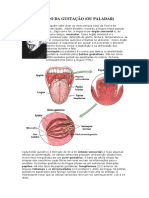
Você também pode gostar
- Perfume e personalidade: neurociência aplicada à perfumariaNo EverandPerfume e personalidade: neurociência aplicada à perfumariaAinda não há avaliações
- A GustaçãoDocumento5 páginasA GustaçãoGeovane SantiagoAinda não há avaliações
- Análise SensorialDocumento4 páginasAnálise SensorialMayza MarquesAinda não há avaliações
- A gastronomia brasileira oitocentista: um estudo lexical dos ingredientes nas obras "Cozinheiro imperial" e "Cozinheiro nacional"No EverandA gastronomia brasileira oitocentista: um estudo lexical dos ingredientes nas obras "Cozinheiro imperial" e "Cozinheiro nacional"Nota: 5 de 5 estrelas5/5 (1)
- Os 4 Sabores Primários da LínguaDocumento6 páginasOs 4 Sabores Primários da LínguaPriscila MendesAinda não há avaliações
- Processos Psicológicos Básicos (PPB) - Olfato + PaladarDocumento16 páginasProcessos Psicológicos Básicos (PPB) - Olfato + Paladarsephiroth1208Ainda não há avaliações
- Sentido Gustativo e OlfativoDocumento6 páginasSentido Gustativo e OlfativoCaroline SantanaAinda não há avaliações
- Trabalho Paladar ProntoDocumento17 páginasTrabalho Paladar Prontonaycg50% (2)
- Fisiologia da GustaçãoDocumento12 páginasFisiologia da GustaçãoGabriel MartinsAinda não há avaliações
- SentidosDocumento8 páginasSentidosIsabelle CeruttiAinda não há avaliações
- 09 Sentido Gustativo OlfativoDocumento6 páginas09 Sentido Gustativo OlfativoDaniel Delani100% (1)
- O sentido da gustaçãoDocumento17 páginasO sentido da gustaçãolilianemachadoAinda não há avaliações
- AULA 12 - Aparelho GustativoDocumento19 páginasAULA 12 - Aparelho GustativoErimilson SicundaAinda não há avaliações
- Fisiologia do paladar e olfatoDocumento2 páginasFisiologia do paladar e olfatoJulia GoicoecheaAinda não há avaliações
- Apg 9 - Vias Especias (Olfato, Paladar e Audição) .Documento16 páginasApg 9 - Vias Especias (Olfato, Paladar e Audição) .Anita DamkeAinda não há avaliações
- Diferença de ReceptoresDocumento3 páginasDiferença de ReceptoresIsaac AlencarAinda não há avaliações
- Aula Paladar Olfato e TatoDocumento33 páginasAula Paladar Olfato e TatoOdilon Calian50% (2)
- Orgãos Do Sentido CompletoDocumento14 páginasOrgãos Do Sentido CompletoValdir FerreiraAinda não há avaliações
- Mecanismo de Percepção de GostoDocumento6 páginasMecanismo de Percepção de GostoDaniela RodriguesAinda não há avaliações
- Sistema Gustativo PDFDocumento33 páginasSistema Gustativo PDFVinicius CorrêaAinda não há avaliações
- Os Orgãos Do Sentido e Seus Mecanismo de FuncionamentoDocumento39 páginasOs Orgãos Do Sentido e Seus Mecanismo de FuncionamentoTaizi RodriguesAinda não há avaliações
- O sentido da gustação e como o cérebro interpreta os sabores dos alimentosDocumento3 páginasO sentido da gustação e como o cérebro interpreta os sabores dos alimentosCastigo VissaiAinda não há avaliações
- AULA 4 - Aná Lise SensorialDocumento31 páginasAULA 4 - Aná Lise SensorialManu FilmesAinda não há avaliações
- Fisiologia - 05 - Órgãos Dos Sentidos - Gustação e OlfaçãoDocumento31 páginasFisiologia - 05 - Órgãos Dos Sentidos - Gustação e OlfaçãoMatheusAinda não há avaliações
- Estomatognático e sentidos químicosDocumento4 páginasEstomatognático e sentidos químicosFúlvia NicollyAinda não há avaliações
- O PaladarDocumento16 páginasO PaladarMuvane CanzerAinda não há avaliações
- Comer Com Todos So SentidosDocumento3 páginasComer Com Todos So SentidosFred MacedoAinda não há avaliações
- Diferença de Receptores IsaacDocumento3 páginasDiferença de Receptores IsaacIsaac AlencarAinda não há avaliações
- OlfatoDocumento20 páginasOlfatoJailaneCostaAinda não há avaliações
- Gostos básicos da análise sensorial de alimentosDocumento20 páginasGostos básicos da análise sensorial de alimentosFelipe Younan100% (1)
- Olfato e GustaçãoDocumento8 páginasOlfato e GustaçãoLusianny ChavesAinda não há avaliações
- Receptores gustativos e olfatoriosDocumento4 páginasReceptores gustativos e olfatoriosTainar VieiraAinda não há avaliações
- A Relevância Metabólica Dos Receptores de SaborDocumento5 páginasA Relevância Metabólica Dos Receptores de SaborPaulo CamargoAinda não há avaliações
- O gosto e sua importânciaDocumento20 páginasO gosto e sua importânciaÉlisson Alex Da SilvaAinda não há avaliações
- Sistema SensorialDocumento26 páginasSistema SensorialDario Cipriano Silverio100% (1)
- Gosto, sabor e paladar: conceitos e percepção sensorialDocumento8 páginasGosto, sabor e paladar: conceitos e percepção sensorialLara Gonçalves MaiaAinda não há avaliações
- Olfato e GustaçãoDocumento1 páginaOlfato e GustaçãoSabrina MagriniAinda não há avaliações
- Os 5 sentidosDocumento2 páginasOs 5 sentidosEsperanca Zanabo AzaleAinda não há avaliações
- Ciencias 6oano C Maa 24072020.docxDocumento3 páginasCiencias 6oano C Maa 24072020.docxEdna LopesAinda não há avaliações
- PALADARDocumento13 páginasPALADARLara PassosAinda não há avaliações
- Quimioreceptores da degustaçãoDocumento10 páginasQuimioreceptores da degustaçãoTainá PestanaAinda não há avaliações
- O Paladar e seus ReceptoresDocumento4 páginasO Paladar e seus ReceptoresEmiliane Martins100% (1)
- 2tics .docx (1)Documento5 páginas2tics .docx (1)Beatriz GoulartAinda não há avaliações
- Gustação e Olfação - EC2019Documento53 páginasGustação e Olfação - EC2019Lucas GustavoAinda não há avaliações
- 5 - Sentidos Quimicos, Olfação e GustaçãoDocumento6 páginas5 - Sentidos Quimicos, Olfação e GustaçãoRafael LopesAinda não há avaliações
- Sensorial Relatorio 2 - Threshold de PercepçãoDocumento10 páginasSensorial Relatorio 2 - Threshold de PercepçãoPaulo Ricardo LosAinda não há avaliações
- Uma Lista de Todos Os Sentidos HumanosDocumento4 páginasUma Lista de Todos Os Sentidos HumanosScribdTranslationsAinda não há avaliações
- Os sentidos e a percepção ambiental nos vertebradosDocumento14 páginasOs sentidos e a percepção ambiental nos vertebradosThais Mello100% (5)
- Resumo Da Lingua Sabor e PapilasDocumento2 páginasResumo Da Lingua Sabor e PapilasMarilay ZacAinda não há avaliações
- Como funcionam os sentidos do paladar e olfatoDocumento11 páginasComo funcionam os sentidos do paladar e olfatoIgor HenriqueAinda não há avaliações
- Arcan WorkDocumento7 páginasArcan WorkFélix AntónioAinda não há avaliações
- Artigo Paladar - Super InteressanteDocumento3 páginasArtigo Paladar - Super InteressanteSarinha MotaAinda não há avaliações
- Os 4 sabores primários captados pelas papilas linguaisDocumento2 páginasOs 4 sabores primários captados pelas papilas linguaisjanainaAinda não há avaliações
- Aula - Receptores SensoriaisDocumento27 páginasAula - Receptores Sensoriaisandersonformiga100% (1)
- Quimiorrecepção - Luiza e EduardoDocumento47 páginasQuimiorrecepção - Luiza e EduardoRoberto SantosAinda não há avaliações
- Apostila Técnicas de Degustação 2023Documento12 páginasApostila Técnicas de Degustação 2023luis.juridicoscAinda não há avaliações
- Aula 04 - Fisio I 2023Documento85 páginasAula 04 - Fisio I 2023Krain SanMeloAinda não há avaliações
- PaladarmeuseminarioDocumento21 páginasPaladarmeuseminarioTaynan GonzagaAinda não há avaliações
- Slide 2Documento6 páginasSlide 2Jan CarlosAinda não há avaliações
- Sistemas de defesa do organismoDocumento14 páginasSistemas de defesa do organismoTatiana PetroffAinda não há avaliações
- Apoptose e NecroseDocumento50 páginasApoptose e NecroseLucas AquinoAinda não há avaliações
- Classificação Dos Seres Vivos PDFDocumento3 páginasClassificação Dos Seres Vivos PDFEfwatAinda não há avaliações
- Etiologia Das Más Oclusôes: Perspectiva Clínica (Parte III) - Fissuras LabiopalatinasDocumento8 páginasEtiologia Das Más Oclusôes: Perspectiva Clínica (Parte III) - Fissuras LabiopalatinasmarianaAinda não há avaliações
- Aula 04.05Documento5 páginasAula 04.05Cris PereiraAinda não há avaliações
- Introdução A Embriologia AnimalDocumento12 páginasIntrodução A Embriologia AnimalANA BEATRIZ MESQUITA DE QUEIROZAinda não há avaliações
- 2017 - UfsmDocumento24 páginas2017 - UfsmTio Leo OliveiraAinda não há avaliações
- Apostila Nuce Ssa1Documento346 páginasApostila Nuce Ssa1RONALD RIBEIRO DOS SANTOSAinda não há avaliações
- Evolução: Lamarck e DarwinDocumento10 páginasEvolução: Lamarck e DarwinJéssica Layne100% (1)
- historepfiles201808RESUMO TECIDO MUSCULAR PDFDocumento4 páginashistorepfiles201808RESUMO TECIDO MUSCULAR PDFDiana Fernandes MirandaAinda não há avaliações
- Descrevendo fertilização e implantação embrionáriaDocumento8 páginasDescrevendo fertilização e implantação embrionáriaRenan VillacaAinda não há avaliações
- Relatório de Aula Prática - Biologia IIDocumento12 páginasRelatório de Aula Prática - Biologia IIBianca Bezerra PiresAinda não há avaliações
- Hematologia RevisãoDocumento2 páginasHematologia RevisãoAndrea MarceleAinda não há avaliações
- Gabarito Simulado Discursivo 01 - COMU 2024Documento2 páginasGabarito Simulado Discursivo 01 - COMU 2024Rafaela fonsecaAinda não há avaliações
- 2 Série - Engenharia GenéticaDocumento29 páginas2 Série - Engenharia GenéticaRafaela PrandoAinda não há avaliações
- Detecção de Coronavírus em Paciente com Teste PCR NegativoDocumento1 páginaDetecção de Coronavírus em Paciente com Teste PCR NegativoMelissa ManhãesAinda não há avaliações
- DNA e Replicação Do DNADocumento4 páginasDNA e Replicação Do DNAGabriel Ferreira100% (1)
- Atividade Darwin LamarckDocumento2 páginasAtividade Darwin LamarckzedroguinhadobairroAinda não há avaliações
- Doença HereditáriaDocumento3 páginasDoença Hereditáriacasa dokameAinda não há avaliações
- Resumo (Medicina Veterinária)Documento8 páginasResumo (Medicina Veterinária)Júlia FreitasAinda não há avaliações
- TCC Emanuelle de Oliveira SouzaDocumento95 páginasTCC Emanuelle de Oliveira SouzaJean SousaAinda não há avaliações
- Cariótipo, mutações e doenças hereditáriasDocumento8 páginasCariótipo, mutações e doenças hereditáriasSamara PereiraAinda não há avaliações
- Proteômica Uma Introdução Aos Métodos e AplicaçõesDocumento11 páginasProteômica Uma Introdução Aos Métodos e AplicaçõesMAGNUS7 BRAinda não há avaliações
- Bioinformática 1Documento12 páginasBioinformática 1TitosAinda não há avaliações
- Lista de Exercícios de Biologia MolecularDocumento2 páginasLista de Exercícios de Biologia MolecularAnderson MatheusAinda não há avaliações
- Biotecnologia III - Técnicas de engenharia genéticaDocumento22 páginasBiotecnologia III - Técnicas de engenharia genéticaJúlia Rigueira HotteAinda não há avaliações
- Caracterização das cianobactérias fotossintéticasDocumento35 páginasCaracterização das cianobactérias fotossintéticasLeonardo LealAinda não há avaliações
- Tecido Conjuntivo: Características e FunçõesDocumento57 páginasTecido Conjuntivo: Características e FunçõesZe NinguenAinda não há avaliações
- Reprodução Das Plantas AtividadesDocumento4 páginasReprodução Das Plantas AtividadesKaela Braga100% (1)
- CADERNO - CNST e MST - @XANDAOENEMTRIDocumento25 páginasCADERNO - CNST e MST - @XANDAOENEMTRIlyvia mariaAinda não há avaliações
- Treinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNo EverandTreinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNota: 4.5 de 5 estrelas4.5/5 (169)
- E-TRAP: entrevista diagnóstica para transtornos de personalidadeNo EverandE-TRAP: entrevista diagnóstica para transtornos de personalidadeNota: 5 de 5 estrelas5/5 (3)
- Educação ambiental: A formação do sujeito ecológicoNo EverandEducação ambiental: A formação do sujeito ecológicoAinda não há avaliações
- Psicoterapia breve: Abordagem sistematizada de situações de criseNo EverandPsicoterapia breve: Abordagem sistematizada de situações de criseNota: 4.5 de 5 estrelas4.5/5 (8)
- Equilíbrio hormonal _ Recupere equilíbrio hormonal, libido, sono e emagreça já!No EverandEquilíbrio hormonal _ Recupere equilíbrio hormonal, libido, sono e emagreça já!Nota: 5 de 5 estrelas5/5 (2)
- Neuromarketing: Como a neurociência aliada ao design pode aumentar o engajamento e a influência sobre os consumidoresNo EverandNeuromarketing: Como a neurociência aliada ao design pode aumentar o engajamento e a influência sobre os consumidoresNota: 5 de 5 estrelas5/5 (7)
- Biomecânica Básica dos Exercícios: Membros InferioresNo EverandBiomecânica Básica dos Exercícios: Membros InferioresNota: 3.5 de 5 estrelas3.5/5 (8)
- Fundamentos Em Cirurgia OralNo EverandFundamentos Em Cirurgia OralNota: 5 de 5 estrelas5/5 (1)
- Ensino de leitura para pessoas com autismoNo EverandEnsino de leitura para pessoas com autismoNota: 4.5 de 5 estrelas4.5/5 (5)
- Uma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNo EverandUma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNota: 4 de 5 estrelas4/5 (13)
- Parasitologia 1: Helmintos de Interesse MédicoNo EverandParasitologia 1: Helmintos de Interesse MédicoAinda não há avaliações
- Atlas de anatomia: Imagens humanas e veterináriasNo EverandAtlas de anatomia: Imagens humanas e veterináriasNota: 4.5 de 5 estrelas4.5/5 (2)
- Emoções: Introdução à psicologia das emoções. De Darwin à neurociência, o que são as emoções e como elas funcionamNo EverandEmoções: Introdução à psicologia das emoções. De Darwin à neurociência, o que são as emoções e como elas funcionamNota: 5 de 5 estrelas5/5 (4)
- Entendendo o Desenvolvimento Infantil: Contribuições das Neurociências e o Papel das Relações Afetivas para Pais e EducadoresNo EverandEntendendo o Desenvolvimento Infantil: Contribuições das Neurociências e o Papel das Relações Afetivas para Pais e EducadoresNota: 5 de 5 estrelas5/5 (4)
- O mundo interior do trauma: Defesas arquetípicas do espírito pessoalNo EverandO mundo interior do trauma: Defesas arquetípicas do espírito pessoalNota: 5 de 5 estrelas5/5 (1)
- Serviços Ecossistêmicos e Planejamento Urbano: A Natureza a Favor do Desenvolvimento Sustentável das CidadesNo EverandServiços Ecossistêmicos e Planejamento Urbano: A Natureza a Favor do Desenvolvimento Sustentável das CidadesAinda não há avaliações
- O tratamento psicanalítico de crianças autistas: Diálogo com múltiplas experiênciasNo EverandO tratamento psicanalítico de crianças autistas: Diálogo com múltiplas experiênciasAinda não há avaliações
- Transtornos Alimentares e NeurociênciaNo EverandTranstornos Alimentares e NeurociênciaAinda não há avaliações