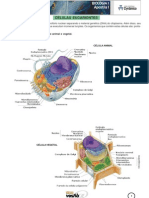
Você também pode gostar
- Metabolismo de LipídeosDocumento5 páginasMetabolismo de LipídeosMarcela Lima de BritoAinda não há avaliações
- 2023 Exercícios - Degradação de Lipídios-Corpos Cetônicos GabaritoDocumento6 páginas2023 Exercícios - Degradação de Lipídios-Corpos Cetônicos GabaritoCybervetWandersonAinda não há avaliações
- Seu metabolismo é incrível: E é fácil compreender como funcionaNo EverandSeu metabolismo é incrível: E é fácil compreender como funcionaAinda não há avaliações
- Metabolismo dos Glicídios: Digestão, Absorção e CirculaçãoDocumento4 páginasMetabolismo dos Glicídios: Digestão, Absorção e CirculaçãoMariana BarbosaAinda não há avaliações
- Dieta Cetogénica: Recetas Rápidas Altas En Grasa Para Restablecer Su Cuerpo Y Vivir Una Vida SaludableNo EverandDieta Cetogénica: Recetas Rápidas Altas En Grasa Para Restablecer Su Cuerpo Y Vivir Una Vida SaludableNota: 5 de 5 estrelas5/5 (1)
- Síntese de lipídeos e esteróidesDocumento3 páginasSíntese de lipídeos e esteróidesMary FreitasAinda não há avaliações
- Digestão e metabolismo dos macronutrientesDocumento13 páginasDigestão e metabolismo dos macronutrientesNayla LimaAinda não há avaliações
- Metabolismo de LipídeosDocumento14 páginasMetabolismo de LipídeosSarah Livramento ZampirolliAinda não há avaliações
- Bioquimica Digestao e Absorcao Dos LipidiosDocumento5 páginasBioquimica Digestao e Absorcao Dos LipidiosdalilaAinda não há avaliações
- Resolução Lista BioquimicaDocumento16 páginasResolução Lista BioquimicaVitor SouzaAinda não há avaliações
- Resumos de BioquimicaDocumento42 páginasResumos de BioquimicaAna Carolina Soares EstevesAinda não há avaliações
- Bioquimica - Digestão e Absorção Dos LipídiosDocumento5 páginasBioquimica - Digestão e Absorção Dos LipídiosRafael Rudá Coelho100% (1)
- Metabolismo Dos LipídiosDocumento5 páginasMetabolismo Dos LipídiosMariana BarbosaAinda não há avaliações
- Bioquímica: Aula 3Documento16 páginasBioquímica: Aula 3Tuani de MirandaAinda não há avaliações
- Metabolismo de CarboidratosDocumento15 páginasMetabolismo de Carboidratosdani_eleine0% (1)
- Metabolismo de lipídios e corpos cetônicosDocumento21 páginasMetabolismo de lipídios e corpos cetônicosDácia ArrudaAinda não há avaliações
- Bioquimica Prova 1 PDFDocumento10 páginasBioquimica Prova 1 PDFESTHER DE MELLO ANTONIO100% (1)
- Metabolismo de LipídeosDocumento8 páginasMetabolismo de LipídeosSabrina XavierAinda não há avaliações
- Resumo Bioquimica Metabólica 1Documento5 páginasResumo Bioquimica Metabólica 1Rafaela BossleAinda não há avaliações
- Resumo - Integração MetabólicaDocumento18 páginasResumo - Integração MetabólicamuradihachichuanAinda não há avaliações
- Lipogênese: como o excesso de carboidratos é armazenado como gorduraDocumento2 páginasLipogênese: como o excesso de carboidratos é armazenado como gorduraMarina De Souza Bastos100% (1)
- Metabolismo dos carboidratos e lipídiosDocumento4 páginasMetabolismo dos carboidratos e lipídiosNetto LacerdaAinda não há avaliações
- Estudo Dirigido I Bioq Med IIDocumento3 páginasEstudo Dirigido I Bioq Med IIArielle MoraesAinda não há avaliações
- Processo de lipogênese e biossíntese de ácidos graxosDocumento5 páginasProcesso de lipogênese e biossíntese de ácidos graxosKAROLINE FAVORETO100% (1)
- Resumo Biologia Celular e MolecularDocumento21 páginasResumo Biologia Celular e Molecularcarolina cardosoAinda não há avaliações
- Lipogênese e armazenamento de TAGDocumento14 páginasLipogênese e armazenamento de TAGPedro Henrique Almeida de BritoAinda não há avaliações
- Síntese de Ácidos GraxosDocumento9 páginasSíntese de Ácidos Graxosfernandoribeirojr98Ainda não há avaliações
- Catabolismo de Ácidos GraxosDocumento8 páginasCatabolismo de Ácidos Graxosjoão vitor fioratti de oliveiraAinda não há avaliações
- A GlicóliseDocumento4 páginasA GlicóliseAna Paula AzevedoAinda não há avaliações
- Apostila I - Bioquímica IIIDocumento17 páginasApostila I - Bioquímica IIIArtur QuiniAinda não há avaliações
- A GlicóliseDocumento4 páginasA GlicóliseAna Paula AzevedoAinda não há avaliações
- Metabolismo de lipídeos: β-oxidação, corpos cetônicos e sínteseDocumento20 páginasMetabolismo de lipídeos: β-oxidação, corpos cetônicos e sínteseVanderval SIlva de OliveiraAinda não há avaliações
- Microsoft Word - Estudo Dirigido - Bloco LipídiosDocumento4 páginasMicrosoft Word - Estudo Dirigido - Bloco LipídiosFelipe MagalhaesAinda não há avaliações
- Lipólise - Degradação dos Ácidos GraxosDocumento6 páginasLipólise - Degradação dos Ácidos GraxosIvone Pires PortoAinda não há avaliações
- 005 Metabolismo de Lipídios (Catabolismo)Documento10 páginas005 Metabolismo de Lipídios (Catabolismo)Izabella Trapia0% (1)
- Metabolismo de LipídiosDocumento16 páginasMetabolismo de LipídiosDanielle RodriguesAinda não há avaliações
- Resumo Bioenergética-RESUMAODocumento4 páginasResumo Bioenergética-RESUMAOVivian Schommer PenhaAinda não há avaliações
- Glicose: digestão, glicólise e regulação dos níveis no sangueDocumento8 páginasGlicose: digestão, glicólise e regulação dos níveis no sanguelaviAinda não há avaliações
- Bioquimica Prova 3Documento7 páginasBioquimica Prova 3LuanaAinda não há avaliações
- Beta oxidação: rota metabólica essencial para a quebra de ácidos graxosDocumento16 páginasBeta oxidação: rota metabólica essencial para a quebra de ácidos graxoskahionz100% (2)
- Bioquímica - Ciclo Alimentado e JejumDocumento9 páginasBioquímica - Ciclo Alimentado e JejumPedro CintraAinda não há avaliações
- Aula 9 - Integração MetabólicaDocumento6 páginasAula 9 - Integração MetabólicaArielle MoraesAinda não há avaliações
- 5 Metabolismo de Lipídios e Aminoácidos e Integração MetabólicaDocumento66 páginas5 Metabolismo de Lipídios e Aminoácidos e Integração MetabólicarilixgsimululadorAinda não há avaliações
- Capítulo 17Documento8 páginasCapítulo 17Davi SousaAinda não há avaliações
- Metabolismo de Lipídeos IQSC-USPDocumento53 páginasMetabolismo de Lipídeos IQSC-USPEvany Priscila100% (1)
- Aula 9 Metabolismo Dos LipídiosDocumento44 páginasAula 9 Metabolismo Dos Lipídiosaracnomarques100% (2)
- oxidacao_beta (1)Documento8 páginasoxidacao_beta (1)Monica AlexandreAinda não há avaliações
- Ciclo de Krebs: oxidação e síntese de ATPDocumento3 páginasCiclo de Krebs: oxidação e síntese de ATPmchamorra100% (1)
- Metabolismo de Carboidratos - Resumo de EscritoDocumento14 páginasMetabolismo de Carboidratos - Resumo de EscritoGualberto Júnior100% (1)
- Lipideos 3Documento42 páginasLipideos 3Diéssica AlvesAinda não há avaliações
- BIOQUIMICADocumento7 páginasBIOQUIMICAWellerson SardiAinda não há avaliações
- RevisãoDocumento6 páginasRevisãoe236610Ainda não há avaliações
- Trabalho BMDocumento3 páginasTrabalho BMraquel farinhaAinda não há avaliações
- Ciclo de Krebs e Efeito WarburgDocumento18 páginasCiclo de Krebs e Efeito WarburgCarolina Almeida100% (1)
- Metabolismo dos Carboidratos: Digestão e Vias MetabólicasDocumento4 páginasMetabolismo dos Carboidratos: Digestão e Vias MetabólicasBia NunesAinda não há avaliações
- Oxidação Dos Ácidos GraxosDocumento5 páginasOxidação Dos Ácidos GraxosBruna NevesAinda não há avaliações
- Metabolismo de Lipídeos Aula 2Documento26 páginasMetabolismo de Lipídeos Aula 2Brunno PinhoAinda não há avaliações
- Metabolismo Dos Carboidratos, Respiração Celular e DesnutriçãoDocumento31 páginasMetabolismo Dos Carboidratos, Respiração Celular e DesnutriçãoLaila LongenAinda não há avaliações
- Resumo de PatologiaDocumento12 páginasResumo de Patologiaapi-3797560100% (11)
- Hemograma completo com leucocitose e neutrofilia leveDocumento1 páginaHemograma completo com leucocitose e neutrofilia leveGilberto MeloAinda não há avaliações
- Morte celular: tipos de necroseDocumento51 páginasMorte celular: tipos de necroseRaylane SilvaasAinda não há avaliações
- Estudo Dirigido Junções CelularesDocumento2 páginasEstudo Dirigido Junções CelularesLarinha VideiraAinda não há avaliações
- Imunogenetica Cap. 13Documento12 páginasImunogenetica Cap. 13meus livrosAinda não há avaliações
- Organelas Celulares: Estruturas do CitoplasmaDocumento31 páginasOrganelas Celulares: Estruturas do CitoplasmaBruno Maria Oliveira100% (1)
- CÉLULASDocumento2 páginasCÉLULASJP CarvalhoAinda não há avaliações
- Aula 2 - Transporte Entre Membranas Biológicas 2017Documento77 páginasAula 2 - Transporte Entre Membranas Biológicas 2017kenyaengAinda não há avaliações
- Bases Científicas Do Treinamento de Hipertrofia PDFDocumento193 páginasBases Científicas Do Treinamento de Hipertrofia PDFGuilherme N100% (11)
- Atividades Sobre Celula em WordDocumento43 páginasAtividades Sobre Celula em WordVanusa SilvaAinda não há avaliações
- Files AP. 1 Biologia 1Documento25 páginasFiles AP. 1 Biologia 1talles123Ainda não há avaliações
- Citologia e Membrana PlasmáticaDocumento31 páginasCitologia e Membrana PlasmáticaMarcelaAinda não há avaliações
- Manual Aluno Bio 10 SantillanaDocumento256 páginasManual Aluno Bio 10 SantillanaBárbara Machado100% (3)
- TeoriaCitoplasma PDFDocumento9 páginasTeoriaCitoplasma PDFShofia SantosAinda não há avaliações
- Ebook Manual Pratico Do EstagiarioDocumento93 páginasEbook Manual Pratico Do EstagiarioanagabrielagorskibortolottoAinda não há avaliações
- Resultado1955806 20230804230721Documento13 páginasResultado1955806 20230804230721rerisson caioAinda não há avaliações
- Policitemia Vera GuiaDocumento10 páginasPolicitemia Vera GuiaJoquebede RodriguesAinda não há avaliações
- Apostila 1.1 de BiologiaDocumento5 páginasApostila 1.1 de BiologiaMaria EduardaAinda não há avaliações
- Biologia Celular: Conceitos e EstruturaDocumento28 páginasBiologia Celular: Conceitos e EstruturaRaquel MarquesAinda não há avaliações
- Estudo APG 02 - Celulas ..Documento10 páginasEstudo APG 02 - Celulas ..Sergio PorcherAinda não há avaliações
- Atividade Sobre Citoplasma - Citosol e Citoesqueleto - 1º Ano EDocumento1 páginaAtividade Sobre Citoplasma - Citosol e Citoesqueleto - 1º Ano EAntonino LopesAinda não há avaliações
- Bomba de sódio e potássioDocumento2 páginasBomba de sódio e potássioLais FernandesAinda não há avaliações
- Observação do Ciclo Celular em Células VegetaisDocumento4 páginasObservação do Ciclo Celular em Células VegetaisafgrhtjykuliçopAinda não há avaliações
- Respiração Aeróbica e AnaeróbicaDocumento3 páginasRespiração Aeróbica e AnaeróbicaDanny SilvaAinda não há avaliações
- Captura de Ecrã 2021-11-09 À(s) 19.16.26Documento1 páginaCaptura de Ecrã 2021-11-09 À(s) 19.16.26Mariana TymchyshynAinda não há avaliações
- Aula CONTROLE DA EXPRESSAO GENICADocumento24 páginasAula CONTROLE DA EXPRESSAO GENICAVERONICA MOREIRA FERREIRAAinda não há avaliações
- Ficha Formativa - TransportesDocumento3 páginasFicha Formativa - TransportesElisaAinda não há avaliações
- Proteínas 2 2Documento57 páginasProteínas 2 2Isabela RochaAinda não há avaliações
- Anatomia Das Plantas de ESAU PDFDocumento43 páginasAnatomia Das Plantas de ESAU PDFgustavotmartins63% (8)
- Aula 07 - Regulação Da Glicose e DiabetesDocumento20 páginasAula 07 - Regulação Da Glicose e DiabetesVictor HigorAinda não há avaliações
- Este não é mais um livro de dieta: O novo e libertador estilo de vida alimentar para saúde e boa forma que derruba o conceito de dietasNo EverandEste não é mais um livro de dieta: O novo e libertador estilo de vida alimentar para saúde e boa forma que derruba o conceito de dietasNota: 4.5 de 5 estrelas4.5/5 (30)
- Hipertrofia Muscular: da Nutrição à SuplementaçãoNo EverandHipertrofia Muscular: da Nutrição à SuplementaçãoAinda não há avaliações
- Cozinha Vegana: 80 Receitas Práticas, Saudáveis E SaborosasNo EverandCozinha Vegana: 80 Receitas Práticas, Saudáveis E SaborosasAinda não há avaliações
- DIETA DASH PARA INICIANTES: Obtenha uma vida saudável perdendo peso de forma rápida, saudável e equilibrada. Reduzir a pressão arterial, inchaço abdominal e inflamação corporalNo EverandDIETA DASH PARA INICIANTES: Obtenha uma vida saudável perdendo peso de forma rápida, saudável e equilibrada. Reduzir a pressão arterial, inchaço abdominal e inflamação corporalNota: 4 de 5 estrelas4/5 (14)
- Detox de 10 dias: Como os sucos verdes limpam o seu organismo e emagrecemNo EverandDetox de 10 dias: Como os sucos verdes limpam o seu organismo e emagrecemNota: 2 de 5 estrelas2/5 (1)
- A alquimia dos sabores: a culinária funcional: Edição revista e atualizadaNo EverandA alquimia dos sabores: a culinária funcional: Edição revista e atualizadaAinda não há avaliações
- Dieta Para Gordura no Fígado Em português/ Diet For Fat In The Liver In PortugueseNo EverandDieta Para Gordura no Fígado Em português/ Diet For Fat In The Liver In PortugueseNota: 5 de 5 estrelas5/5 (1)
- Uma dieta além da moda: Uma abordagem científica para a perda de peso e a manutenção da sáudeNo EverandUma dieta além da moda: Uma abordagem científica para a perda de peso e a manutenção da sáudeNota: 4.5 de 5 estrelas4.5/5 (4)
- 21 HiperSucos: para saúde física, mental e longevidadeNo Everand21 HiperSucos: para saúde física, mental e longevidadeAinda não há avaliações
- Saúde Intestinal, Limpeza Hepática, Fadiga Adrenal, Depressão, Ansiedade, Perda De Peso E Pele Sadia: Cure Sua Mente, Corpo E Hormônios Com Dieta, Nutrição, Medicina Natural E Terapias HolísticasNo EverandSaúde Intestinal, Limpeza Hepática, Fadiga Adrenal, Depressão, Ansiedade, Perda De Peso E Pele Sadia: Cure Sua Mente, Corpo E Hormônios Com Dieta, Nutrição, Medicina Natural E Terapias HolísticasNota: 5 de 5 estrelas5/5 (7)
- Sem glúten: Pães, tortas e bolos sem restriçõesNo EverandSem glúten: Pães, tortas e bolos sem restriçõesNota: 5 de 5 estrelas5/5 (3)
- Dieta Anti-inflamatória EstratégicaNo EverandDieta Anti-inflamatória EstratégicaNota: 5 de 5 estrelas5/5 (2)
- Livro de Receitas da Dieta Anti-inflamatória de 7 Dias E Plano Fácil de Reduzir a Inflamação Em portuguêsNo EverandLivro de Receitas da Dieta Anti-inflamatória de 7 Dias E Plano Fácil de Reduzir a Inflamação Em portuguêsNota: 5 de 5 estrelas5/5 (4)
- Treino de Peso Corporal Em português/ Body Weight Training In PortugueseNo EverandTreino de Peso Corporal Em português/ Body Weight Training In PortugueseAinda não há avaliações
- Emagrecer: Perda de Peso: Livro de Receitas Diet para Vigilantes de Peso (Para Aqueles Que Gostam De Dietas)No EverandEmagrecer: Perda de Peso: Livro de Receitas Diet para Vigilantes de Peso (Para Aqueles Que Gostam De Dietas)Ainda não há avaliações
- Dieta Mediterrânea - A Ciência E A Arte Da Dieta Mediterrânea: Um Guia Completo Para Iniciantes Para Queimar Gordura E Alcançar Uma Perda De Peso PermanenteNo EverandDieta Mediterrânea - A Ciência E A Arte Da Dieta Mediterrânea: Um Guia Completo Para Iniciantes Para Queimar Gordura E Alcançar Uma Perda De Peso PermanenteAinda não há avaliações
- Mais energia: Dieta para aumentar a resistência e dar mais energiaNo EverandMais energia: Dieta para aumentar a resistência e dar mais energiaAinda não há avaliações
- Estimulação Precoce e Assessoria: Discursos em CenaNo EverandEstimulação Precoce e Assessoria: Discursos em CenaAinda não há avaliações
- Mente sã, corpo são: O poder de uma mente saudávelNo EverandMente sã, corpo são: O poder de uma mente saudávelNota: 5 de 5 estrelas5/5 (3)
- Simetria Molecular III: Série Didática para o apoio a formação de professores de QuímicaNo EverandSimetria Molecular III: Série Didática para o apoio a formação de professores de QuímicaAinda não há avaliações