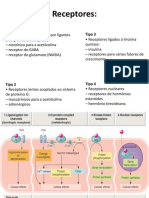
Você também pode gostar
- Aula 9Documento23 páginasAula 9Diogo CostaAinda não há avaliações
- 1.2 Mecanismo de Transdução HormonalDocumento3 páginas1.2 Mecanismo de Transdução HormonalJaqueline SantosAinda não há avaliações
- 029 Biossinalizacao 009Documento41 páginas029 Biossinalizacao 009Manuela M. Lins100% (1)
- BQI 241 OkDocumento13 páginasBQI 241 OkCaio AugustusAinda não há avaliações
- Aula 6 e 7 Sinalização Celular 2019Documento33 páginasAula 6 e 7 Sinalização Celular 2019Dinis SousaAinda não há avaliações
- Aula RECEPTORES DE MEMBRANA E VIAS DE SINALIZAÇÃO s2Documento2 páginasAula RECEPTORES DE MEMBRANA E VIAS DE SINALIZAÇÃO s2JúliaAinda não há avaliações
- Aula 4-Bio Cel e MolDocumento42 páginasAula 4-Bio Cel e MolFrancielli Valerio de OliveiraAinda não há avaliações
- Aula 6 - Transdução de Sinais BiológicosDocumento12 páginasAula 6 - Transdução de Sinais BiológicosJailton MagalhãesAinda não há avaliações
- Aula Sinalização I PDFDocumento60 páginasAula Sinalização I PDFLuizCarlosVilarAinda não há avaliações
- 19.1 Sinalizaçao CelularDocumento6 páginas19.1 Sinalizaçao CelularAna CristinaAinda não há avaliações
- Sinalização CelularDocumento6 páginasSinalização CelularChristianAinda não há avaliações
- Comunicacao Quimica Entre Celulas 100407Documento25 páginasComunicacao Quimica Entre Celulas 100407Profa Raíza MeloAinda não há avaliações
- Receptores Ligados A CanaisDocumento5 páginasReceptores Ligados A Canaiskarinecampos10100% (1)
- 4a Aula SinapseDocumento83 páginas4a Aula SinapseGabriel Messias100% (1)
- Aula 01 - COMUNICAÇÃO, SINALIZAÇÃO CELULAR E HOMEOSTASIADocumento7 páginasAula 01 - COMUNICAÇÃO, SINALIZAÇÃO CELULAR E HOMEOSTASIADaniel MunizAinda não há avaliações
- BiossinalizaçãoDocumento19 páginasBiossinalizaçãoSergio Correa FernandesAinda não há avaliações
- As Epatas Da ApoptoseDocumento16 páginasAs Epatas Da ApoptosejoanaAinda não há avaliações
- Bioquímica II - Resumo IDocumento10 páginasBioquímica II - Resumo ISabrina AraujoAinda não há avaliações
- Trabalho Sistema Seg MensageirosDocumento11 páginasTrabalho Sistema Seg MensageirosScarlett DuarteAinda não há avaliações
- BiocelDocumento10 páginasBiocelPatriciaAinda não há avaliações
- 2 Aula FarmadinamicaDocumento48 páginas2 Aula FarmadinamicaEduardo CostaAinda não há avaliações
- Proteina G PDFDocumento8 páginasProteina G PDFAna CristinaAinda não há avaliações
- PORTFÓLIODocumento43 páginasPORTFÓLIOJaqueline AlvesAinda não há avaliações
- Biossinalização - Beatriz MayumiDocumento3 páginasBiossinalização - Beatriz Mayumibe.mayumi.2010Ainda não há avaliações
- Sinalização IntracelularDocumento4 páginasSinalização IntracelularAna CristinaAinda não há avaliações
- Bioquimica - Sinalizacao Celular PDFDocumento6 páginasBioquimica - Sinalizacao Celular PDFFernando CastanheiraAinda não há avaliações
- Atividades de BiossinalizaçãoDocumento3 páginasAtividades de Biossinalizaçãobe.mayumi.2010Ainda não há avaliações
- Aula 4 Sinalização Celular 2017 EBDocumento17 páginasAula 4 Sinalização Celular 2017 EBAloy Da CruzAinda não há avaliações
- Vias de SinalizaçãoDocumento4 páginasVias de SinalizaçãoTayse SantosAinda não há avaliações
- SinalizaçãoDocumento4 páginasSinalizaçãomônica conceiçãoAinda não há avaliações
- Sinalizaçao Celular - Nível UniversitárioDocumento93 páginasSinalizaçao Celular - Nível UniversitárioPit100% (5)
- HormonasDocumento31 páginasHormonasxicana83% (6)
- Aula 5 - Receptores Farmacológicos e Sistemas EfetoresDocumento27 páginasAula 5 - Receptores Farmacológicos e Sistemas EfetoresRodolfo Rodrigues Zacarias100% (4)
- Comunicação e Morte Celular ProgramadaDocumento19 páginasComunicação e Morte Celular ProgramadaWellington SoaresAinda não há avaliações
- BVE 3009 - AULA 1-Conceitos de Crescimento e DesenvolvimentoDocumento44 páginasBVE 3009 - AULA 1-Conceitos de Crescimento e DesenvolvimentoLaís VictóriaAinda não há avaliações
- Farmacologia ReceptoresDocumento46 páginasFarmacologia ReceptorescamilleAinda não há avaliações
- Tipos de Sinapses ModulatóriasDocumento5 páginasTipos de Sinapses ModulatóriasMrkinghtgAinda não há avaliações
- 48584Documento27 páginas48584Josyane Aparecida BezerraAinda não há avaliações
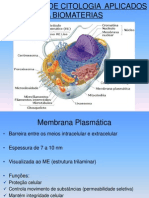
- Conceitos de Citologia Aplicados A BiomateriaisDocumento56 páginasConceitos de Citologia Aplicados A BiomateriaisSthéfany PiresAinda não há avaliações
- Revisão BIO CEL IDocumento27 páginasRevisão BIO CEL IgiovanaAinda não há avaliações
- SinalizaçãoDocumento4 páginasSinalizaçãoMariana PaesAinda não há avaliações
- Mecanismos de Ação Dos Fármacos.Documento6 páginasMecanismos de Ação Dos Fármacos.Gessyka SousaAinda não há avaliações
- Cell Cycle 2017Documento41 páginasCell Cycle 2017João Frederico MusialAinda não há avaliações
- Sinalização Celular Parte 2 - FisiologiaDocumento8 páginasSinalização Celular Parte 2 - FisiologiaGabriella do CarmoAinda não há avaliações
- BiossinalizaçãoDocumento17 páginasBiossinalizaçãoMelissaAlvesAinda não há avaliações
- Aula 11 e 12-CalcioDocumento29 páginasAula 11 e 12-CalcioLouis LFRAinda não há avaliações
- Farmacodinâmica Odontologia I 2023-1 - Parte ReceptoresDocumento58 páginasFarmacodinâmica Odontologia I 2023-1 - Parte ReceptoresJúnia Rosa De SouzaAinda não há avaliações
- CEDERJ-Biologia Celular I - AulaDocumento14 páginasCEDERJ-Biologia Celular I - Aulaapi-3800070Ainda não há avaliações
- Biologia Celular e Genética - Aula 05 - Sinalização CelularDocumento20 páginasBiologia Celular e Genética - Aula 05 - Sinalização CelularDouglas NobreAinda não há avaliações
- CEDERJ Biologia Celular I - Gabarito 13 A 20Documento9 páginasCEDERJ Biologia Celular I - Gabarito 13 A 20api-3800070Ainda não há avaliações
- Aulas 11 e 12Documento39 páginasAulas 11 e 12Diogo CostaAinda não há avaliações
- 02 - Classificação Dos HormôniosDocumento3 páginas02 - Classificação Dos HormôniosLuis OliveiraAinda não há avaliações
- Resposta AdrenérgicosDocumento4 páginasResposta AdrenérgicosManoel Neto100% (1)
- Comunicação e Sinalização CelularDocumento14 páginasComunicação e Sinalização CelularGonçalo MachadoAinda não há avaliações
- 6 - Princípios Da Ação Dos Fármacos 2023Documento31 páginas6 - Princípios Da Ação Dos Fármacos 2023Rayssa SilvaAinda não há avaliações
- Ciclo Celular 2021Documento63 páginasCiclo Celular 2021Ana Livia CamolesiAinda não há avaliações
- BiossinalizaçãoDocumento4 páginasBiossinalizaçãojones100% (1)
- Terapia Gênica com hPEDF via vetores AAVs e Nanopartículas na Asma AlérgicaNo EverandTerapia Gênica com hPEDF via vetores AAVs e Nanopartículas na Asma AlérgicaAinda não há avaliações
- Prolactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoNo EverandProlactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoAinda não há avaliações
- Teste de Conhecimento 2 Genética Forense PDFDocumento3 páginasTeste de Conhecimento 2 Genética Forense PDFGiovanna AraujoAinda não há avaliações
- 2 1001 2a Serie Aula N2 2024Documento15 páginas2 1001 2a Serie Aula N2 2024Lucas BattistiAinda não há avaliações
- Células Imunes SagahDocumento17 páginasCélulas Imunes SagahsogixiAinda não há avaliações
- Aula Teorica 10 - Eudicots - RosideasFabideas - 2017Documento105 páginasAula Teorica 10 - Eudicots - RosideasFabideas - 2017Hermes Sales CamposAinda não há avaliações
- Como Nos Tornamos Humanos (2010)Documento159 páginasComo Nos Tornamos Humanos (2010)luquinhasonceAinda não há avaliações
- 10 Tulipia Academy - Linha Moovy Redutor 2Documento63 páginas10 Tulipia Academy - Linha Moovy Redutor 2Maisa RodriguesAinda não há avaliações
- Atividade AdaptadaDocumento3 páginasAtividade AdaptadaRondiney RodriguesAinda não há avaliações
- InsulinaDocumento1 páginaInsulinaNathan Henrique Rodrigues LimaAinda não há avaliações
- Embriologia IDocumento30 páginasEmbriologia ILindsei BitencourtAinda não há avaliações
- Cultura Celular AnimalDocumento2 páginasCultura Celular AnimalKayo PaivaAinda não há avaliações
- Codigo GenéticoDocumento25 páginasCodigo GenéticoJoão RibeiroAinda não há avaliações
- SÉRIE MELIPONICULTURA #08 Meliponicultura Perguntas Mais Frequentes Sobre As Abelhas Sem FerrãoDocumento49 páginasSÉRIE MELIPONICULTURA #08 Meliponicultura Perguntas Mais Frequentes Sobre As Abelhas Sem FerrãoCleiton BernardoAinda não há avaliações
- Insect Metamorphosis From Natural History To Regulation of Development and Evolution (Xavier Belles) (Z-Lib - Org) - CompressedDocumento294 páginasInsect Metamorphosis From Natural History To Regulation of Development and Evolution (Xavier Belles) (Z-Lib - Org) - CompressedRenata PereiraAinda não há avaliações
- Periquito Catarina - 2014Documento6 páginasPeriquito Catarina - 2014Edson França RodriguesAinda não há avaliações
- Teste 1 2021 MJAM VADocumento7 páginasTeste 1 2021 MJAM VAarm.bioAinda não há avaliações
- Resumao AlgasDocumento1 páginaResumao AlgasRayssa RosaAinda não há avaliações
- Síndrome de DownDocumento4 páginasSíndrome de DownManuAinda não há avaliações
- Aula 2 - Leis de Mendel - FabioDocumento26 páginasAula 2 - Leis de Mendel - FabioTaiane GamaAinda não há avaliações
- Trypanosoma CruziDocumento17 páginasTrypanosoma CruziEduarda GomesAinda não há avaliações
- Embriologia e Anatomia Do Sistema Reprodutor FemininoDocumento49 páginasEmbriologia e Anatomia Do Sistema Reprodutor FemininoHansAinda não há avaliações
- Exercicios Evolucao - Principios de Hardy-WeinbergDocumento3 páginasExercicios Evolucao - Principios de Hardy-WeinbergSheyla Souza DareAinda não há avaliações
- Preparacao para ExameDocumento5 páginasPreparacao para ExamePuto SantanaAinda não há avaliações
- Simulado Biologia CasafDocumento16 páginasSimulado Biologia CasafRaissa SantiagoAinda não há avaliações
- Aula 9 - Herança MonogênicaDocumento32 páginasAula 9 - Herança MonogênicaEVERTON100% (1)
- Testes Intermedios Compilacao Biologia 11º AnoDocumento40 páginasTestes Intermedios Compilacao Biologia 11º AnoMargarida AnjosAinda não há avaliações
- O Que É A Comunicação Celular e para Que Serve?Documento6 páginasO Que É A Comunicação Celular e para Que Serve?EnzoAinda não há avaliações
- R - T - Leandro Mattos SantosDocumento83 páginasR - T - Leandro Mattos SantosManuel AugustoAinda não há avaliações
- Aula - Estudo Dirigido - Micro, Parasito e Epidemiologia - 2023 - 1Documento34 páginasAula - Estudo Dirigido - Micro, Parasito e Epidemiologia - 2023 - 1Yasmin AzeredoAinda não há avaliações
- Captura de Tela 2023-05-03 À(s) 08.05.51Documento32 páginasCaptura de Tela 2023-05-03 À(s) 08.05.5105armadas-basesAinda não há avaliações
- MARTELOTA - Exercícios Sobre Gerativismo Do Livro Manual de LinguísticaDocumento5 páginasMARTELOTA - Exercícios Sobre Gerativismo Do Livro Manual de LinguísticaSAMANTHA LAISA SIMAO CUNHAAinda não há avaliações