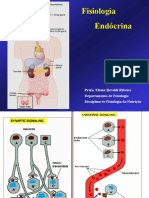
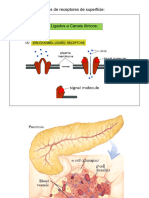
Você também pode gostar
- Aula 4-Bio Cel e MolDocumento42 páginasAula 4-Bio Cel e MolFrancielli Valerio de OliveiraAinda não há avaliações
- Aula 5 - Sinalização CelularDocumento37 páginasAula 5 - Sinalização CelularCamila HerculanoAinda não há avaliações
- 029 Biossinalizacao 009Documento41 páginas029 Biossinalizacao 009Manuela M. Lins100% (1)
- Aula 9Documento23 páginasAula 9Diogo CostaAinda não há avaliações
- Receptores Ligados A CanaisDocumento5 páginasReceptores Ligados A Canaiskarinecampos10100% (1)
- Aula RECEPTORES DE MEMBRANA E VIAS DE SINALIZAÇÃO s2Documento2 páginasAula RECEPTORES DE MEMBRANA E VIAS DE SINALIZAÇÃO s2JúliaAinda não há avaliações
- Proteina G PDFDocumento8 páginasProteina G PDFAna CristinaAinda não há avaliações
- Aula Sinalização I PDFDocumento60 páginasAula Sinalização I PDFLuizCarlosVilarAinda não há avaliações
- Cell Cycle 2017Documento41 páginasCell Cycle 2017João Frederico MusialAinda não há avaliações
- Farmacodinâmica Odontologia I 2023-1 - Parte ReceptoresDocumento58 páginasFarmacodinâmica Odontologia I 2023-1 - Parte ReceptoresJúnia Rosa De SouzaAinda não há avaliações
- Aula 6 - Transdução de Sinais BiológicosDocumento12 páginasAula 6 - Transdução de Sinais BiológicosJailton MagalhãesAinda não há avaliações
- 1.2 Mecanismo de Transdução HormonalDocumento3 páginas1.2 Mecanismo de Transdução HormonalJaqueline SantosAinda não há avaliações
- Sinalização CelularDocumento6 páginasSinalização CelularChristianAinda não há avaliações
- Bioquímica - MonitoriaDocumento7 páginasBioquímica - MonitoriaNyna PaulaAinda não há avaliações
- Sinalização Celular 2018.2Documento38 páginasSinalização Celular 2018.2Baba BabaAinda não há avaliações
- Mecanismos de Ação Dos Fármacos.Documento6 páginasMecanismos de Ação Dos Fármacos.Gessyka SousaAinda não há avaliações
- BQI 241 OkDocumento13 páginasBQI 241 OkCaio AugustusAinda não há avaliações
- A DigestãoDocumento61 páginasA DigestãoRomário WelterAinda não há avaliações
- NeurotransmissoresDocumento63 páginasNeurotransmissoresJessica LeeAinda não há avaliações
- 4a Aula SinapseDocumento83 páginas4a Aula SinapseGabriel Messias100% (1)
- Expressão GênicaDocumento6 páginasExpressão GênicaLaysla ManuelyAinda não há avaliações
- Aula 11 e 12-CalcioDocumento29 páginasAula 11 e 12-CalcioLouis LFRAinda não há avaliações
- Sinalização Celular Parte 2 - FisiologiaDocumento8 páginasSinalização Celular Parte 2 - FisiologiaGabriella do CarmoAinda não há avaliações
- Aulas 11 e 12Documento39 páginasAulas 11 e 12Diogo CostaAinda não há avaliações
- G-Protein Lecture HUDocumento42 páginasG-Protein Lecture HUIsa SilvaAinda não há avaliações
- Resposta AdrenérgicosDocumento4 páginasResposta AdrenérgicosManoel Neto100% (1)
- Aulão para 1 Prova de MBI 100Documento22 páginasAulão para 1 Prova de MBI 100Rodrigo Antônio Pires VieiraAinda não há avaliações
- Resumo BCMDocumento4 páginasResumo BCMJoão Guilherme Lira AcioliAinda não há avaliações
- Trabalho Sistema Seg MensageirosDocumento11 páginasTrabalho Sistema Seg MensageirosScarlett DuarteAinda não há avaliações
- Aula 4 Sinapses e NeurotransmissoresDocumento65 páginasAula 4 Sinapses e NeurotransmissoresRonneo LúcioAinda não há avaliações
- RegulacaodometabolismoDocumento24 páginasRegulacaodometabolismoJoao Paulo Fagundes LafetaAinda não há avaliações
- Balanço Ca, P e MGDocumento4 páginasBalanço Ca, P e MGjenniferAinda não há avaliações
- MineralocorticóidesDocumento13 páginasMineralocorticóidesEvander AngeloAinda não há avaliações
- BioqDocumento30 páginasBioqMichelle BarcellosAinda não há avaliações
- 2016 Mestrado Prova-Escrita Completa Com-Gabarito-3 PDFDocumento13 páginas2016 Mestrado Prova-Escrita Completa Com-Gabarito-3 PDFFernando Henrique DeslockAinda não há avaliações
- Sinalização CelularDocumento3 páginasSinalização CelularGiovanna GuimarãesAinda não há avaliações
- Receptores Adrenérgicos Edição FinalDocumento23 páginasReceptores Adrenérgicos Edição Finaljosé santosAinda não há avaliações
- Bioquímica II - Resumo IDocumento10 páginasBioquímica II - Resumo ISabrina AraujoAinda não há avaliações
- Aula 4 - Metabolismo de AADocumento29 páginasAula 4 - Metabolismo de AASoniaAinda não há avaliações
- Processos Bioenergéticos e NeoglicogêneseDocumento50 páginasProcessos Bioenergéticos e NeoglicogêneseyvignoloAinda não há avaliações
- Aula Mecanismo EndocDocumento52 páginasAula Mecanismo EndocabresserAinda não há avaliações
- Expressão GênicaDocumento7 páginasExpressão GênicaAna CristinaAinda não há avaliações
- Aula9 Expressao Genica EucariotosDocumento63 páginasAula9 Expressao Genica EucariotosCoordenação Ciências BiológicasAinda não há avaliações
- Aprendizado e MemoriaDocumento22 páginasAprendizado e MemoriaMarcusVeniciusAinda não há avaliações
- 01 - Respiração Celular Anaeróbica e AeróbicaDocumento74 páginas01 - Respiração Celular Anaeróbica e AeróbicaJoão Victor MolinaAinda não há avaliações
- Ciclo Do Ácido CítricoDocumento8 páginasCiclo Do Ácido CítricoFernanda Eder FerreiraAinda não há avaliações
- Sinalizaà à o 2 Presencial Fono - PPTX - 20240407 - 110933 - 0000Documento26 páginasSinalizaà à o 2 Presencial Fono - PPTX - 20240407 - 110933 - 0000raquel2005freitasAinda não há avaliações
- Aula 5Documento4 páginasAula 5Marcos CarvalhoAinda não há avaliações
- BiossinalizaçãoDocumento19 páginasBiossinalizaçãoSergio Correa FernandesAinda não há avaliações
- Glicólise - BQM IIDocumento5 páginasGlicólise - BQM IIJAQUELINE AGUIARAinda não há avaliações
- Ciclo Celular 2021Documento63 páginasCiclo Celular 2021Ana Livia CamolesiAinda não há avaliações
- B OxidacaoDocumento49 páginasB OxidacaoMariana Gonçalves MacedoAinda não há avaliações
- Farmacodinâmica de MedicamentosDocumento40 páginasFarmacodinâmica de MedicamentosJosimar JúniorAinda não há avaliações
- Introdução Ao Metabolismo - Metabolismo de Carboidrato - UBCDocumento101 páginasIntrodução Ao Metabolismo - Metabolismo de Carboidrato - UBCLeiliane RibeiroAinda não há avaliações
- 19.1 Sinalizaçao CelularDocumento6 páginas19.1 Sinalizaçao CelularAna CristinaAinda não há avaliações
- Metabolismo de CarboidratosDocumento8 páginasMetabolismo de CarboidratosGabriel FerreiraAinda não há avaliações
- Sebenta Biocel BDLDocumento210 páginasSebenta Biocel BDLcsanchezp72Ainda não há avaliações
- Comunicacao Quimica Entre Celulas 100407Documento25 páginasComunicacao Quimica Entre Celulas 100407Profa Raíza MeloAinda não há avaliações
- Terapia Gênica com hPEDF via vetores AAVs e Nanopartículas na Asma AlérgicaNo EverandTerapia Gênica com hPEDF via vetores AAVs e Nanopartículas na Asma AlérgicaAinda não há avaliações
- 0-Bases de QuímicaDocumento32 páginas0-Bases de QuímicaDinis SousaAinda não há avaliações
- Aula 5 - Sinalização Celular 2019Documento29 páginasAula 5 - Sinalização Celular 2019Dinis SousaAinda não há avaliações
- Aula3-Núcleo e CitoesqueletoDocumento45 páginasAula3-Núcleo e CitoesqueletoDiana VicenteAinda não há avaliações
- Aula 4 - Organelos Celulares - BTDocumento118 páginasAula 4 - Organelos Celulares - BTDiana VicenteAinda não há avaliações
- Aula 2 - Membrana PlasmáticaDocumento38 páginasAula 2 - Membrana PlasmáticaDiana VicenteAinda não há avaliações
- Aula 1 - Introdução À BiologiaDocumento92 páginasAula 1 - Introdução À BiologiaDinis SousaAinda não há avaliações
- Elenco Bluebook 805Documento652 páginasElenco Bluebook 805damminsonAinda não há avaliações
- Melhore A Resistência Com AlongamentoDocumento2 páginasMelhore A Resistência Com AlongamentoRac A BruxaAinda não há avaliações
- Programação Web - Conheça As Principais LinguagensDocumento6 páginasProgramação Web - Conheça As Principais LinguagensIvan Cortez CtzAinda não há avaliações
- Atividade - Prática Penal - Relaxamento de Prisão em Flagrante - Nathália Matos Lima - OdtDocumento3 páginasAtividade - Prática Penal - Relaxamento de Prisão em Flagrante - Nathália Matos Lima - OdtNathalia LimaAinda não há avaliações
- I3 Termo de Referencia EaiDocumento8 páginasI3 Termo de Referencia Eaifelipe thiago senaAinda não há avaliações
- Identificar As Áreas Cerebrais de Controle RespiratórioDocumento16 páginasIdentificar As Áreas Cerebrais de Controle RespiratórioGustavo CamposAinda não há avaliações
- Introducao A Biblioteconomia e Ciencia Da InformacaoDocumento5 páginasIntroducao A Biblioteconomia e Ciencia Da InformacaoWeilaAinda não há avaliações
- José Pastore - Custos Dos Acidentes de TrabalhoDocumento5 páginasJosé Pastore - Custos Dos Acidentes de TrabalhoAnonymous MjVTyRTGdqAinda não há avaliações
- Apostila 6º Ano - Egito Antigo - Esta Atividade Faz Parte Das Atividades Liberadas Do Material de DivulgaçãoDocumento4 páginasApostila 6º Ano - Egito Antigo - Esta Atividade Faz Parte Das Atividades Liberadas Do Material de DivulgaçãoAndre SilvaAinda não há avaliações
- Comp - Mario Cristiano Trindade ManuelDocumento2 páginasComp - Mario Cristiano Trindade ManuelJussara PereiraAinda não há avaliações
- RESOLUÇÃO CFO-196, de 29 de Janeiro de 2019Documento2 páginasRESOLUÇÃO CFO-196, de 29 de Janeiro de 2019isa057Ainda não há avaliações
- DINÂMICADocumento5 páginasDINÂMICALucas AppeltAinda não há avaliações
- Modulo IV Via em CurvaDocumento45 páginasModulo IV Via em CurvafilipeAinda não há avaliações
- AutoCad - Como Organizar Blocos e Inserir em Qualquer ProjetoDocumento7 páginasAutoCad - Como Organizar Blocos e Inserir em Qualquer Projetoleandro_rafaelAinda não há avaliações
- Questionário para Exame de TomografiaDocumento1 páginaQuestionário para Exame de TomografiaCei SAÚDEAinda não há avaliações
- Apostila - História e Peculiaridades Do Método Pilates, Uma Abordagem InterdisciplinarDocumento44 páginasApostila - História e Peculiaridades Do Método Pilates, Uma Abordagem InterdisciplinarPedala FormosaAinda não há avaliações
- Bloco 4 - 240110 - 230503Documento9 páginasBloco 4 - 240110 - 230503marianaAinda não há avaliações
- 8 TransitividadeDocumento4 páginas8 TransitividadeAdrianeAinda não há avaliações
- Receituario Bolas BerlimDocumento16 páginasReceituario Bolas BerlimfisiocaciaAinda não há avaliações
- A Sentença CristãDocumento1 páginaA Sentença CristãmarcinhakrAinda não há avaliações
- Sequencia DidaticaDocumento5 páginasSequencia DidaticaIcaro Benevides100% (2)
- Infraçoes Disciplinares e Processo DisciplinarDocumento156 páginasInfraçoes Disciplinares e Processo Disciplinartiagoacofe2015Ainda não há avaliações
- Melanie PIII E01Documento31 páginasMelanie PIII E01Melanie FonsecaAinda não há avaliações
- Proc - Sejus - Tortura Psma IIDocumento202 páginasProc - Sejus - Tortura Psma IIAmafavv AmafavvAinda não há avaliações
- Histologia Prática 3 - Prova 2Documento8 páginasHistologia Prática 3 - Prova 2carolinacantoniAinda não há avaliações
- Temas 2 BimestreDocumento6 páginasTemas 2 BimestreBruno FragaAinda não há avaliações
- Ensinar Não É Transferir ConhecimentoDocumento1 páginaEnsinar Não É Transferir ConhecimentokmartinskmsAinda não há avaliações
- RDC - 25 - 2001 - Comercializacao de Equipamentos UsadosDocumento3 páginasRDC - 25 - 2001 - Comercializacao de Equipamentos UsadosGustavo BastosAinda não há avaliações
- Planner 30dias Top5Documento8 páginasPlanner 30dias Top5Madah RolimAinda não há avaliações
- Simulado de Administração e Cálculo de MedicamentosDocumento8 páginasSimulado de Administração e Cálculo de MedicamentosJennifer LimaAinda não há avaliações