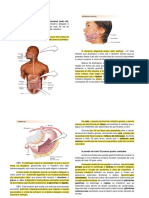
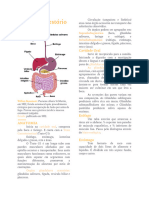
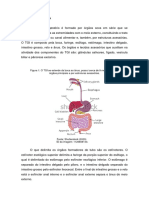
Você também pode gostar
- Periodo CompostoDocumento34 páginasPeriodo CompostoRefúgio SeguroAinda não há avaliações
- Tratamentos de Ultima Geracao Francisco Borrello RadiestesiaDocumento12 páginasTratamentos de Ultima Geracao Francisco Borrello RadiestesiaJaine Silva100% (1)
- Paee - Plano de Atendimento Educacional Especializado-José Carlos Gonçalves (2) FinalizadoDocumento10 páginasPaee - Plano de Atendimento Educacional Especializado-José Carlos Gonçalves (2) FinalizadoAndriele Monteiro100% (2)
- Solomons ResolvidoDocumento142 páginasSolomons ResolvidoAmanda Ramos87% (15)
- Metodologia Científica (SLIDES DA AULA)Documento67 páginasMetodologia Científica (SLIDES DA AULA)Arts Begbie100% (2)
- Apostila DisbioseDocumento26 páginasApostila DisbioseJean Cloudy PimentelAinda não há avaliações
- Parábolas de Jesus - 2022Documento146 páginasParábolas de Jesus - 2022Hilmar CesarAinda não há avaliações
- Chapeuzinho Vermelho - PerraultDocumento103 páginasChapeuzinho Vermelho - PerraultSâmia Sousa100% (1)
- Remédios Caseiros para Desparasitar Gatos - PeritoAnimalDocumento7 páginasRemédios Caseiros para Desparasitar Gatos - PeritoAnimalMarcos Antonio da SilvaAinda não há avaliações
- Arte Abstrata 22 AgostoDocumento55 páginasArte Abstrata 22 Agostovickymsantos17Ainda não há avaliações
- Lição Completa Tradição e SabedoriaDocumento194 páginasLição Completa Tradição e SabedoriaDaniel GomesAinda não há avaliações
- Pop Uenc 002Documento12 páginasPop Uenc 002Juliana Coelho100% (1)
- Up6 Parte Ii (Digestivo)Documento105 páginasUp6 Parte Ii (Digestivo)Cláudia FonsecaAinda não há avaliações
- Fisiologia Gastrintestinal Motilidade GastrointestinalDocumento7 páginasFisiologia Gastrintestinal Motilidade GastrointestinalMatheus MoraesAinda não há avaliações
- Sistema GastrointestinalDocumento12 páginasSistema GastrointestinalDoce MulherAinda não há avaliações
- Camadas IntestinoDocumento2 páginasCamadas IntestinoMatheus R10Ainda não há avaliações
- Camadas Intestino PDFDocumento2 páginasCamadas Intestino PDFMatheus R10Ainda não há avaliações
- MOTILIDADEDocumento22 páginasMOTILIDADEJhuly SantanaAinda não há avaliações
- Conteúdos Relativos Ao Teste de Língua Inglesa.Documento56 páginasConteúdos Relativos Ao Teste de Língua Inglesa.DaviAinda não há avaliações
- Histologia E Embriologia: Adriana Dalpicolli RodriguesDocumento26 páginasHistologia E Embriologia: Adriana Dalpicolli Rodriguesjanainaaparecidadejesus22Ainda não há avaliações
- DigestórioDocumento103 páginasDigestórioAndré RainanAinda não há avaliações
- Resumo - Sistema DigestórioDocumento19 páginasResumo - Sistema DigestórioVinícius AlvesAinda não há avaliações
- Apresentação Completa Do Sistema DigestivoDocumento27 páginasApresentação Completa Do Sistema DigestivoFavorite FilmesAinda não há avaliações
- Resumo Do Sistema DigestórioDocumento37 páginasResumo Do Sistema DigestórioighormtAinda não há avaliações
- 2 - Sistema DigestórioDocumento6 páginas2 - Sistema Digestórioisinha6952Ainda não há avaliações
- Digestorio 4Documento22 páginasDigestorio 4Joyce SouzaAinda não há avaliações
- PP Instestino Delgado AnatomofisiologiaDocumento15 páginasPP Instestino Delgado AnatomofisiologiaVasco DiasAinda não há avaliações
- Sistema DigestórioDocumento59 páginasSistema DigestórioLollaAinda não há avaliações
- Anatomia Do Sistema DigestórioDocumento74 páginasAnatomia Do Sistema DigestórioBianca FranciscoAinda não há avaliações
- Ciencias Morfofuncionais Sist. Digestório, Edógrino e RenalDocumento30 páginasCiencias Morfofuncionais Sist. Digestório, Edógrino e RenalMaríliah GiudiceAinda não há avaliações
- Fisiologia Do Sistema Digestório.P3Documento24 páginasFisiologia Do Sistema Digestório.P3thatsmarianaAinda não há avaliações
- Sistema Digestório Órgãos e Funções Do Trato GastrintestinalDocumento14 páginasSistema Digestório Órgãos e Funções Do Trato GastrintestinalelizafernandanutriAinda não há avaliações
- Sistema DigestórioDocumento13 páginasSistema DigestórioIsadora RodriguesAinda não há avaliações
- Documento Ciencias MolecularesDocumento5 páginasDocumento Ciencias MolecularesTallita SilvaAinda não há avaliações
- Aula 9 - Fisiologia Do Sistema GastrointestinalDocumento18 páginasAula 9 - Fisiologia Do Sistema GastrointestinalElizael GoncalvesAinda não há avaliações
- Fascículo de Biologia (2022-2023) - 8 ClasseDocumento27 páginasFascículo de Biologia (2022-2023) - 8 ClasseAntónio LízuaAinda não há avaliações
- Anatomia e Histologia Do Sistema Digestório Alto e BaixoDocumento7 páginasAnatomia e Histologia Do Sistema Digestório Alto e BaixoLuís EduardoAinda não há avaliações
- Estudo Dirigido - Problema 2 ASE 5Documento28 páginasEstudo Dirigido - Problema 2 ASE 5ANDRESSA DA CRUZ GURGELAinda não há avaliações
- Sistema Digestório - CadernoDocumento8 páginasSistema Digestório - CadernoduhchiamoleraAinda não há avaliações
- 6º Etapa - Morfo Módulo1 PDFDocumento77 páginas6º Etapa - Morfo Módulo1 PDFMatheus ParavizoAinda não há avaliações
- TEMA 7 - Fisiologia Do Sistema DigestórioDocumento62 páginasTEMA 7 - Fisiologia Do Sistema DigestórioMaryB.HbrandtAinda não há avaliações
- Resumo de Sistema DigestórioDocumento8 páginasResumo de Sistema DigestóriolaiskomatiAinda não há avaliações
- Histologia - Esôfago e EstômagoDocumento3 páginasHistologia - Esôfago e EstômagogabrieltavaresAinda não há avaliações
- Sistema Digestório Roteiro PersonalizadoDocumento9 páginasSistema Digestório Roteiro Personalizadoaninha.cddiasAinda não há avaliações
- Anatomia Do Sistema DigestórioDocumento81 páginasAnatomia Do Sistema DigestórioGeovanna Uchoa FurtadoAinda não há avaliações
- P2 - LucianoDocumento16 páginasP2 - LucianoWashington DuarteAinda não há avaliações
- DiuréticosDocumento9 páginasDiuréticosFernand TananAinda não há avaliações
- Estudo Dirigido 3Documento6 páginasEstudo Dirigido 3Elisa MartinsAinda não há avaliações
- 02 Gastro MotilidadeDocumento8 páginas02 Gastro MotilidadeJoana VindeirinhoAinda não há avaliações
- Processo de Disgestão e Absorção de Nutrientes.Documento42 páginasProcesso de Disgestão e Absorção de Nutrientes.Ariane Azevedo PinheiroAinda não há avaliações
- Ficha Informativa 2Documento5 páginasFicha Informativa 2Anonymous dPwFX8TIBMAinda não há avaliações
- Histologia - Intestino DelgadoDocumento4 páginasHistologia - Intestino DelgadoPaula OliveiraAinda não há avaliações
- Anatomia Do Sistema DigestórioDocumento113 páginasAnatomia Do Sistema DigestórioMilena MarinhoAinda não há avaliações
- Aula 05Documento14 páginasAula 05Vitoria SalesAinda não há avaliações
- Sistema Digestório ResumoDocumento9 páginasSistema Digestório ResumoLuíza BarrosAinda não há avaliações
- Sistema DigestórioDocumento34 páginasSistema DigestórioMaria Stella FernandesAinda não há avaliações
- 1 Explique Ou Esquematize A Organização Anatômica Do Sistema DigestórioDocumento3 páginas1 Explique Ou Esquematize A Organização Anatômica Do Sistema Digestórioesfbarra barraAinda não há avaliações
- Anatomorfofisiologia Un. 1 - DigestórioDocumento57 páginasAnatomorfofisiologia Un. 1 - DigestórioLucas EspindolaAinda não há avaliações
- Resumão 2 ProvaDocumento9 páginasResumão 2 ProvaMaria IsabelAinda não há avaliações
- Abordagem Morfofuncional Do Sistema Digestorio PDFDocumento45 páginasAbordagem Morfofuncional Do Sistema Digestorio PDFwfkamAinda não há avaliações
- PeritoneuDocumento11 páginasPeritoneuDuarte RufinoAinda não há avaliações
- Aulas de Anatomia e Fisiologia PadraoDocumento258 páginasAulas de Anatomia e Fisiologia PadraorachidonaboranaboraAinda não há avaliações
- Aula - DigestórioDocumento52 páginasAula - Digestórioehh noisAinda não há avaliações
- Aparelho Digestivo - Wikipédia, A Enciclopédia LivreDocumento6 páginasAparelho Digestivo - Wikipédia, A Enciclopédia LivreDionísio GuidioneAinda não há avaliações
- Leitura DigestórioDocumento13 páginasLeitura Digestórioallan orengoAinda não há avaliações
- Fisiologia 5Documento11 páginasFisiologia 5Aline KreiAinda não há avaliações
- Resumo - Sistemas Digestivo, Urinário e ReprodutorDocumento12 páginasResumo - Sistemas Digestivo, Urinário e ReprodutorThayane MoreiraAinda não há avaliações
- Apostila Sobre o Sistema DigestórioDocumento10 páginasApostila Sobre o Sistema DigestórioVeronicalg20Ainda não há avaliações
- Mod 3 P07Documento15 páginasMod 3 P07Rayane rodrigues dos santosAinda não há avaliações
- Ciência Da NutriçãoDocumento12 páginasCiência Da NutriçãoRayane rodrigues dos santosAinda não há avaliações
- Mod 03 P05Documento16 páginasMod 03 P05Rayane rodrigues dos santosAinda não há avaliações
- Tuturial 4 - Fisioterapia, Saúde e FuncionalidadeDocumento17 páginasTuturial 4 - Fisioterapia, Saúde e FuncionalidadeRayane rodrigues dos santosAinda não há avaliações
- Ciência Da NutriçãoDocumento8 páginasCiência Da NutriçãoRayane rodrigues dos santosAinda não há avaliações
- Relatório PECDocumento5 páginasRelatório PECRayane rodrigues dos santosAinda não há avaliações
- Documento 14Documento3 páginasDocumento 14Rayane rodrigues dos santosAinda não há avaliações
- Fechamento 05Documento10 páginasFechamento 05Rayane rodrigues dos santosAinda não há avaliações
- Fechamento 04Documento10 páginasFechamento 04Rayane rodrigues dos santosAinda não há avaliações
- Fechamento 1Documento11 páginasFechamento 1Rayane rodrigues dos santosAinda não há avaliações
- Fechamento P03Documento12 páginasFechamento P03Rayane rodrigues dos santosAinda não há avaliações
- Funções Biologicas Vias Respiratorias SuperioresDocumento5 páginasFunções Biologicas Vias Respiratorias SuperioresRayane rodrigues dos santosAinda não há avaliações
- Relatório Final PECDocumento10 páginasRelatório Final PECRayane rodrigues dos santosAinda não há avaliações
- Aula II - Sistema CardiovascularDocumento4 páginasAula II - Sistema CardiovascularRayane rodrigues dos santosAinda não há avaliações
- Relatório Do MapaDocumento8 páginasRelatório Do MapaRayane rodrigues dos santosAinda não há avaliações
- Pós-Fechamento Problema 01Documento7 páginasPós-Fechamento Problema 01Rayane rodrigues dos santosAinda não há avaliações
- Redação de Educação FísicaDocumento1 páginaRedação de Educação FísicaRayane rodrigues dos santosAinda não há avaliações
- Pedagogia Do Oprimido - Capítulo 1Documento2 páginasPedagogia Do Oprimido - Capítulo 1Rayane rodrigues dos santosAinda não há avaliações
- Hipotálamo e EpitálamoDocumento31 páginasHipotálamo e EpitálamoCamila CarneiroAinda não há avaliações
- Proposta Comercial GtraceDocumento10 páginasProposta Comercial GtraceFernando MachadoAinda não há avaliações
- Telhado Stock House - Tripper CarDocumento6 páginasTelhado Stock House - Tripper CarDavid OliveiraAinda não há avaliações
- Preservado para Um PropósitoDocumento8 páginasPreservado para Um Propósitoalanariana2644Ainda não há avaliações
- Fatura Vivo 10-2023Documento3 páginasFatura Vivo 10-2023Paulo FreitasAinda não há avaliações
- 1.2 Resumo Introdução - AracnídeosDocumento4 páginas1.2 Resumo Introdução - AracnídeosPaloma BaldisseraAinda não há avaliações
- Resolution A.960 (23) TraducaoDocumento11 páginasResolution A.960 (23) TraducaoMarco Antonio ToscanoAinda não há avaliações
- Divina de Jesus Scarpim - Deus Não Existe e Eu Posso Provar PDFDocumento265 páginasDivina de Jesus Scarpim - Deus Não Existe e Eu Posso Provar PDFLuciano de Oliveira0% (1)
- Sistemas Políticos Comparados - ResumosDocumento36 páginasSistemas Políticos Comparados - Resumosgabriela.resendeAinda não há avaliações
- 196-Texto Do Artigo-825-794-10-20170119Documento9 páginas196-Texto Do Artigo-825-794-10-20170119Edmilson Mendes Meireles JuniorAinda não há avaliações
- Back-UPS BR - BZ1500XLBI-BRDocumento3 páginasBack-UPS BR - BZ1500XLBI-BRbruno cominAinda não há avaliações
- SlidesDocumento52 páginasSlidesWillian Ricardo RitterAinda não há avaliações
- Estatística e FiabilidadeDocumento54 páginasEstatística e FiabilidadeAnaAinda não há avaliações
- Magias Divinas Do 4º Círculo Reais de Fato ReaisDocumento11 páginasMagias Divinas Do 4º Círculo Reais de Fato ReaisDiego Silva DrumondAinda não há avaliações
- Material Didatico - Treinamento NR35Documento8 páginasMaterial Didatico - Treinamento NR35FREAKAZOIDEAinda não há avaliações
- O Biogás Na AlemanhaDocumento2 páginasO Biogás Na AlemanhaMatheus RodriguesAinda não há avaliações
- Iguaba Grande - Wikipédia, A Enciclopédia LivreDocumento4 páginasIguaba Grande - Wikipédia, A Enciclopédia LivreJulio Cezar Souza100% (1)
- Resultado - Mãe - João Paulo LemosDocumento1 páginaResultado - Mãe - João Paulo LemosDavidson GuimarãesAinda não há avaliações
- Dodf 043 03-03-2023 IntegraDocumento62 páginasDodf 043 03-03-2023 IntegraMáscaras FaciaisAinda não há avaliações