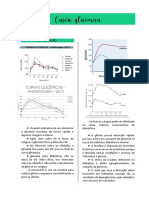
Você também pode gostar
- Prolactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoNo EverandProlactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoAinda não há avaliações
- Metabolismo de CarboidratosDocumento6 páginasMetabolismo de CarboidratosEronAinda não há avaliações
- Glicogênese e Glicogenólise OKDocumento3 páginasGlicogênese e Glicogenólise OKlau24bronzattoAinda não há avaliações
- Biossíntese de Oses e ÓsidosDocumento29 páginasBiossíntese de Oses e ÓsidosÏśä Čhäntrë LïmäAinda não há avaliações
- Form Obj 0Documento33 páginasForm Obj 0Daniele Dantas De Oliveira100% (1)
- 04 Estrutura e Metabolismo Dos Carboidratos - CompressedDocumento50 páginas04 Estrutura e Metabolismo Dos Carboidratos - CompressedMarcos LimaAinda não há avaliações
- Aula03 BioqII-CFBio Glioconeogenese1Documento38 páginasAula03 BioqII-CFBio Glioconeogenese1hide-seekAinda não há avaliações
- Pâncreas EndócrinoDocumento29 páginasPâncreas EndócrinoDennisFrankKondo100% (1)
- Unidade 3 Catabolismo de Carbo Digestão e Absorçao NovoDocumento65 páginasUnidade 3 Catabolismo de Carbo Digestão e Absorçao NovoThais Nunes SouzaAinda não há avaliações
- Metabolismo Do GlicogênioDocumento24 páginasMetabolismo Do GlicogênioDominique SakagamiAinda não há avaliações
- GliconeogeneseDocumento9 páginasGliconeogeneseRayssa CabralAinda não há avaliações
- Aula 04 - GlicogênioDocumento30 páginasAula 04 - GlicogênioVictor HigorAinda não há avaliações
- Regulação Da Glicólise e Diferença Da Hexoquinase e GlicoquinaseDocumento1 páginaRegulação Da Glicólise e Diferença Da Hexoquinase e Glicoquinasejbarb201150% (4)
- Aula 5 - Regulação Do Metabolismo CarboidratosDocumento25 páginasAula 5 - Regulação Do Metabolismo CarboidratosJosé Augusto CeronAinda não há avaliações
- Gliconeogenese e Metabolismo Do GlicogênioDocumento27 páginasGliconeogenese e Metabolismo Do GlicogênioRicardo BorgesAinda não há avaliações
- Glicoólise e Gliconeogenese e GlicogenioDocumento46 páginasGlicoólise e Gliconeogenese e Glicogeniobrunaco2006Ainda não há avaliações
- Aula - Metabolismo Do GlicogênioDocumento16 páginasAula - Metabolismo Do Glicogêniojuliana moraes niloAinda não há avaliações
- Resumos BioquímicaDocumento14 páginasResumos BioquímicaGabrieli HoepersAinda não há avaliações
- Introdução Ao MetabolismoDocumento6 páginasIntrodução Ao MetabolismoLuanaAinda não há avaliações
- Sintese e Degradação Do Glicogênio e CKDocumento33 páginasSintese e Degradação Do Glicogênio e CKwandersom costa de souzaAinda não há avaliações
- HipoglicemiantesDocumento31 páginasHipoglicemiantesvitoriaAinda não há avaliações
- AT14 GliconeogeneseDocumento30 páginasAT14 GliconeogeneseMateus GarciaAinda não há avaliações
- Von Gierke B2Documento23 páginasVon Gierke B2abresserAinda não há avaliações
- Insulina+e+glucagon+2017 2 PDFDocumento12 páginasInsulina+e+glucagon+2017 2 PDFLucas LimaAinda não há avaliações
- CiclosDocumento62 páginasCiclosMJTAinda não há avaliações
- Glicogênese, Glicogenólise e PentosePDocumento4 páginasGlicogênese, Glicogenólise e PentosePVivian Leão VeterináriaAinda não há avaliações
- GliconeogêneseDocumento2 páginasGliconeogênesepatrickdrheonixAinda não há avaliações
- GluconeogeneseDocumento27 páginasGluconeogenesebeatrizasilvapdmaAinda não há avaliações
- Via Glicolítica IVDocumento28 páginasVia Glicolítica IVSabrina AraujoAinda não há avaliações
- Glicolise Bioquimica SlidDocumento41 páginasGlicolise Bioquimica SlidSidvaldo Sid SousaAinda não há avaliações
- Resumo REGULACAO DA GLICOLISEDocumento5 páginasResumo REGULACAO DA GLICOLISEJOCOSO GPLAYAinda não há avaliações
- Aula 2 - Metabolismo Da Glicose (Farmácia 2013)Documento55 páginasAula 2 - Metabolismo Da Glicose (Farmácia 2013)jaqueft5239100% (1)
- Continuação Via Glicolítica 16Documento7 páginasContinuação Via Glicolítica 16Reinaldo RiquetAinda não há avaliações
- 15) NeoglicogêneseDocumento4 páginas15) NeoglicogêneseMary FreitasAinda não há avaliações
- Resumo - GlicóliseDocumento11 páginasResumo - Glicólisemaelson100% (2)
- Glicolise 1Documento14 páginasGlicolise 1zekynhaAinda não há avaliações
- Bioquímica-Fisiológica-Exame 210129 010644Documento7 páginasBioquímica-Fisiológica-Exame 210129 010644Joana Carolina Teixeira RibeiroAinda não há avaliações
- Regulação Do MetabolismoDocumento6 páginasRegulação Do MetabolismoJohn FroesAinda não há avaliações
- Aula 12Documento33 páginasAula 12leonor rochaAinda não há avaliações
- Metabolismo Do GlicognioDocumento15 páginasMetabolismo Do GlicognioDaniel FreireAinda não há avaliações
- Metabolismo Glicognio Regulao2022Documento35 páginasMetabolismo Glicognio Regulao2022amanda abreuAinda não há avaliações
- KerbsDocumento2 páginasKerbsJuliana Dalmas100% (1)
- Glicólise MonitoriaDocumento13 páginasGlicólise MonitoriaLara SalesAinda não há avaliações
- Curva GlicêmicaDocumento4 páginasCurva GlicêmicaRê SilvaAinda não há avaliações
- 2 GlicóliseDocumento39 páginas2 GlicóliseNathan RodriguesAinda não há avaliações
- GliconeogeneseDocumento4 páginasGliconeogeneseAline PereiraAinda não há avaliações
- 11 Glicogenese e GlicogenoliseDocumento23 páginas11 Glicogenese e GlicogenoliseAna Cláudia GomesAinda não há avaliações
- Via GlicolticaDocumento15 páginasVia GlicolticaCamila MendesAinda não há avaliações
- Aula 14 - GluconeogéneseDocumento24 páginasAula 14 - GluconeogéneseaptgaptgAinda não há avaliações
- UntitledDocumento5 páginasUntitledAna RosaAinda não há avaliações
- Moodle2pluginfile - Php611568mod Resourcecontent1SC3ADntese20e20degradação20de20glicogêni 2Documento41 páginasMoodle2pluginfile - Php611568mod Resourcecontent1SC3ADntese20e20degradação20de20glicogêni 2Ana CarolineAinda não há avaliações
- Bioquimica II Estudo Dirigido - Primeira Prova - FARMACIA UFOBDocumento10 páginasBioquimica II Estudo Dirigido - Primeira Prova - FARMACIA UFOBShâmara Stéfany GuimarãesAinda não há avaliações
- BioquimiscDocumento7 páginasBioquimiscJuca Do sabugueiro loucoAinda não há avaliações
- Gliconeogênese e Metabolismo de Glicídios - RespostasDocumento2 páginasGliconeogênese e Metabolismo de Glicídios - RespostasMirella DannaAinda não há avaliações
- Aula Pancreas 1Documento32 páginasAula Pancreas 1silvia pradoAinda não há avaliações
- 15 - Pancreas EndocrinoDocumento33 páginas15 - Pancreas Endocrinokelen.navarroAinda não há avaliações
- AULA 05 - Metabolísmo GlicogênioDocumento36 páginasAULA 05 - Metabolísmo GlicogênioReinaldo Riquet100% (1)
- Aula de Glicogênio Via Pentoses NeoglicogêneseDocumento70 páginasAula de Glicogênio Via Pentoses NeoglicogêneseLaysFitaroniAinda não há avaliações
- Biossintese de Carboidratos BioquímicaDocumento6 páginasBiossintese de Carboidratos BioquímicaMarcela Lima de BritoAinda não há avaliações
- Máquina para Solda MIG/MAG Voonder MM 251Documento36 páginasMáquina para Solda MIG/MAG Voonder MM 251Alexandre S. CorrêaAinda não há avaliações
- Clara Flaksman - Narrativas Relacoes Emaranhados PDFDocumento293 páginasClara Flaksman - Narrativas Relacoes Emaranhados PDFCarolina Freitas Peres100% (2)
- A Degradaã Ã o Dos Solos MILENADocumento7 páginasA Degradaã Ã o Dos Solos MILENAMilena Laureano Lima CardosoAinda não há avaliações
- QueimadurasDocumento8 páginasQueimadurasPaulo Fernando da SilvaAinda não há avaliações
- Conan 40 AnosDocumento4 páginasConan 40 AnosRonan BarrosAinda não há avaliações
- Chat Da LiveDocumento904 páginasChat Da LiveMarcosAinda não há avaliações
- Ranger 82 A 118Documento21 páginasRanger 82 A 118Vilmar Gnoato50% (2)
- Cores de Complexo de CobreDocumento3 páginasCores de Complexo de CobreImerson Mota100% (1)
- (Versão 02) - Conceitos e Técnicas Básicas para Fabricação de Embalagens em MadeiraDocumento22 páginas(Versão 02) - Conceitos e Técnicas Básicas para Fabricação de Embalagens em MadeiraArlindo CrispimAinda não há avaliações
- Textos de Umbanda Extraidos Da InternetDocumento83 páginasTextos de Umbanda Extraidos Da InternetValeria RibeiroAinda não há avaliações
- EXERCICIOS UNIPAZ-PR Dez-2017Documento14 páginasEXERCICIOS UNIPAZ-PR Dez-2017Bruno HenriqueAinda não há avaliações
- Código Irt 2020Documento2 páginasCódigo Irt 2020Sérgio Magalhães100% (1)
- Banner Indigenas 3.0-1Documento1 páginaBanner Indigenas 3.0-1Souza cris LiraAinda não há avaliações
- Lilith e Príapo Na AstrologiaDocumento53 páginasLilith e Príapo Na AstrologiaKarla Mattos VaidyaratnaAinda não há avaliações
- PJF - Aprovaçâo de Projetos 2019Documento84 páginasPJF - Aprovaçâo de Projetos 2019Alexandra RampinelliAinda não há avaliações
- Parafrase PDFDocumento5 páginasParafrase PDFZé SilvaAinda não há avaliações
- Apostila Curso Express Power BIDocumento158 páginasApostila Curso Express Power BIerinaldopaladinoAinda não há avaliações
- Ficha de Português 2º Período - AdapatadaDocumento4 páginasFicha de Português 2º Período - AdapatadaSandra VieiraAinda não há avaliações
- Wayne Mack - A Utilidade Das Escrituras No Aconselhamento PDFDocumento4 páginasWayne Mack - A Utilidade Das Escrituras No Aconselhamento PDFHugo Santos ZicaAinda não há avaliações
- Questões de Enfermagem - Parte 1 @estudajaDocumento118 páginasQuestões de Enfermagem - Parte 1 @estudajaGabi MessiAinda não há avaliações
- Texto 2 (B) Parte II - Lopes e Macedo (2011)Documento39 páginasTexto 2 (B) Parte II - Lopes e Macedo (2011)josephantonivs51960% (1)
- Extração de Amido Batata e Lipidio AmendoimDocumento2 páginasExtração de Amido Batata e Lipidio AmendoimRenataAinda não há avaliações
- Atividade de Semântica IIDocumento5 páginasAtividade de Semântica IISandrinha PereiraAinda não há avaliações
- Cartilha Acessibilidade - Web PDFDocumento130 páginasCartilha Acessibilidade - Web PDFmaiquelveigaAinda não há avaliações
- PROJETO PPSUS Aprovado Comitê de Etica-Jan21-Modificadoabril21Documento31 páginasPROJETO PPSUS Aprovado Comitê de Etica-Jan21-Modificadoabril21fotoseduardaamaralAinda não há avaliações
- Doenças Venosas e ArteriaisDocumento8 páginasDoenças Venosas e ArteriaisWanessa KallyneAinda não há avaliações
- Língua Portuguesa - Agnaldo MartinoDocumento8 páginasLíngua Portuguesa - Agnaldo Martinocamilapati100% (1)
- Apostila de Artes VisuaisDocumento59 páginasApostila de Artes VisuaisDaniele Lamim100% (2)
- Cad C2 Teoria 1serie 20aulas 2bim 2023 PortuguesDocumento4 páginasCad C2 Teoria 1serie 20aulas 2bim 2023 PortuguesFERNANDA FERREIRA DOS SANTOSAinda não há avaliações
- 2p. SAVIANI, N. Saber Escolar, Currículo e Didática - Problemas Da Unidade Conteúdo - Método No Processo Pedagógico.Documento2 páginas2p. SAVIANI, N. Saber Escolar, Currículo e Didática - Problemas Da Unidade Conteúdo - Método No Processo Pedagógico.Marco Túlio Santana dos Reis100% (2)