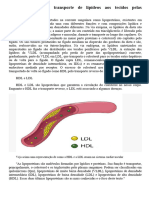
Você também pode gostar
- HDL Colesterol: Transporte de Colesterol E Funções Das HdlsDocumento9 páginasHDL Colesterol: Transporte de Colesterol E Funções Das HdlsSôraya GuedesAinda não há avaliações
- Bases Fisiopatologicas Da Dislipidemia AterogenicaDocumento7 páginasBases Fisiopatologicas Da Dislipidemia AterogenicaPedroAinda não há avaliações
- Bioquimica Atividade de Estudo 2Documento3 páginasBioquimica Atividade de Estudo 2Thaiane KistAinda não há avaliações
- Diretriz Brasileira Sobre Dislipidemias PDFDocumento18 páginasDiretriz Brasileira Sobre Dislipidemias PDFaafernandsAinda não há avaliações
- Aula 2 DislipidemiasDocumento74 páginasAula 2 Dislipidemiaspatricia torkaski100% (1)
- 1586733641-Metabolismo LipidicoDocumento10 páginas1586733641-Metabolismo LipidicoMarcos HenriquesAinda não há avaliações
- Roteiro Do Laboratório Morfofuncional: Curso de MedicinaDocumento30 páginasRoteiro Do Laboratório Morfofuncional: Curso de MedicinabluboarettoAinda não há avaliações
- 4 Aula Final Do Capítulo de ProteínasDocumento3 páginas4 Aula Final Do Capítulo de Proteínaslaliatelie2022Ainda não há avaliações
- Bioquímica Clínica - Perfil Lipídico 2021Documento44 páginasBioquímica Clínica - Perfil Lipídico 2021Karina Felinto0% (1)
- Dislipidemia - ResumoDocumento8 páginasDislipidemia - ResumoEduardo BarretoAinda não há avaliações
- Roteiro Do Laboratório Morfofuncional: Curso de MedicinaDocumento32 páginasRoteiro Do Laboratório Morfofuncional: Curso de MedicinabluboarettoAinda não há avaliações
- Dislipidemia SanarDocumento28 páginasDislipidemia SanarRoberta Violante100% (1)
- ED FiÌ - GadoDocumento5 páginasED FiÌ - GadoRafaela MendesAinda não há avaliações
- Síntese Aula 7 (Lipídios)Documento3 páginasSíntese Aula 7 (Lipídios)Julyana RochaAinda não há avaliações
- Clinica 2 - Prova 2Documento13 páginasClinica 2 - Prova 2VITORIA PEREIRA ALVES MELOAinda não há avaliações
- Seção 2 Unidade 3Documento23 páginasSeção 2 Unidade 3Yam VictorAinda não há avaliações
- Aula - Perfil LipidicoDocumento7 páginasAula - Perfil Lipidicoluansantos1993Ainda não há avaliações
- Cartilha DislipidemiaDocumento12 páginasCartilha DislipidemiaRayane Silva100% (1)
- Interpretação de Exames (Reparado)Documento131 páginasInterpretação de Exames (Reparado)Jefferson Peres de MacedoAinda não há avaliações
- Dislipidemias e Prevenção Da AteroscleroseDocumento9 páginasDislipidemias e Prevenção Da AteroscleroseAlice MirandaAinda não há avaliações
- Como Ocorre o Transporte de Lipídeos Aos Tecidos Pelas Lipoproteínas4Documento2 páginasComo Ocorre o Transporte de Lipídeos Aos Tecidos Pelas Lipoproteínas4Sanderson Samid de Vasconcelos MaiaAinda não há avaliações
- Doseamento de LípidosDocumento4 páginasDoseamento de LípidosAlexandra MarquesAinda não há avaliações
- Aula 2Documento36 páginasAula 2LuisAinda não há avaliações
- DislipidemiasDocumento11 páginasDislipidemiasEvelyn385Ainda não há avaliações
- Lipidios, Apolipos e LipoproteinasDocumento105 páginasLipidios, Apolipos e LipoproteinasMarval Fone100% (1)
- Ebook Gorduras e Saúde CardiovascularDocumento20 páginasEbook Gorduras e Saúde CardiovascularClézia CarvalhoAinda não há avaliações
- Metabolismo Lipídico e Doenças CardiovascularesDocumento31 páginasMetabolismo Lipídico e Doenças CardiovascularesAdarly KrothAinda não há avaliações
- Sistema Digestório - Colesterol PDFDocumento7 páginasSistema Digestório - Colesterol PDFPolliana CastroAinda não há avaliações
- DislipidemiaDocumento14 páginasDislipidemiajoao bernacchiAinda não há avaliações
- Questões ColesterolDocumento2 páginasQuestões ColesterolAndréia LaraAinda não há avaliações
- Bioquímica Clínica de Lipídeos NovoDocumento34 páginasBioquímica Clínica de Lipídeos NovoHatsona MikuAinda não há avaliações
- LipoproteínasDocumento57 páginasLipoproteínasDiego Iglesias100% (1)
- Aula 6 Lipídeos, Lipoproteínas e ApolipoproteínasDocumento50 páginasAula 6 Lipídeos, Lipoproteínas e ApolipoproteínasLídia Rosa100% (1)
- DislipidemiaDocumento17 páginasDislipidemiaAbel Chacanessa de JesusAinda não há avaliações
- Aula - DislipidemiaDocumento22 páginasAula - DislipidemiaallansalvadorpereiraAinda não há avaliações
- LIPÍDIOS Fechamento UC IVDocumento8 páginasLIPÍDIOS Fechamento UC IVLuis Mateus Camelo França MartinsAinda não há avaliações
- Transcrição Dos Áudios 2 Prova de Bioquímica Clínica IDocumento16 páginasTranscrição Dos Áudios 2 Prova de Bioquímica Clínica ICarol Fernandes de Assis100% (1)
- Problema 03 11Documento20 páginasProblema 03 11Ricardo Vitor Mesquita BedettiAinda não há avaliações
- Aula DislipidemiasDocumento4 páginasAula DislipidemiasRayssa LimaAinda não há avaliações
- Consenso Brasileiro Sobre DislipidemiasDocumento19 páginasConsenso Brasileiro Sobre DislipidemiasEtiena MaritanAinda não há avaliações
- Nutrição Nas Doenças Cronicas Não TransmissiveisDocumento20 páginasNutrição Nas Doenças Cronicas Não Transmissiveislidia_barbosaAinda não há avaliações
- Perfil LipídicoDocumento63 páginasPerfil LipídicoJosyane Aparecida Bezerra100% (1)
- RESUMO Metabolismo Das LipoproteínasDocumento7 páginasRESUMO Metabolismo Das LipoproteínasSalomão SandesAinda não há avaliações
- Lipoproteà Nas e Metabolismo Do Colesterol - Nutrià à o 2022-1Documento59 páginasLipoproteà Nas e Metabolismo Do Colesterol - Nutrià à o 2022-1Julia Corsini100% (1)
- Transporte e Armazenamento de LipideosDocumento6 páginasTransporte e Armazenamento de LipideosCamila LopesAinda não há avaliações
- LipidoseDocumento10 páginasLipidoseSérgio HenriqueAinda não há avaliações
- BioquimicaDocumento6 páginasBioquimicakesianadielyAinda não há avaliações
- Cap 7 - Farmacoterapia Das DislipidemiasDocumento3 páginasCap 7 - Farmacoterapia Das DislipidemiasAna Carolyna VargasAinda não há avaliações
- Diretrizes Dislipidemia 2017 - SOCESPDocumento37 páginasDiretrizes Dislipidemia 2017 - SOCESPMônica Amaral LenziAinda não há avaliações
- Relatorio de Dosagem de Triglicerides e As DislipidemiasDocumento11 páginasRelatorio de Dosagem de Triglicerides e As DislipidemiasJohnny Percussionista100% (3)
- DISLIPIDEMIADocumento2 páginasDISLIPIDEMIAbira_tanAinda não há avaliações
- Colesterol TotalDocumento5 páginasColesterol TotalJoice CruzAinda não há avaliações
- Metabolismo Do Colesterol - ResumidoDocumento5 páginasMetabolismo Do Colesterol - ResumidoDianaAinda não há avaliações
- Alterações Do Metabolismo Lipídico No Paciente Com Síndrome MetabólicaDocumento6 páginasAlterações Do Metabolismo Lipídico No Paciente Com Síndrome MetabólicaPablo EduardoAinda não há avaliações
- Bioquímica Clínica - Aula 4 - Perfil LipídicoDocumento26 páginasBioquímica Clínica - Aula 4 - Perfil LipídicoValeskaSena100% (2)
- Questoes BioquimicaDocumento12 páginasQuestoes BioquimicaGleycimara Patricia100% (5)
- Metabolismo de Lipídeos em Ruminantes PDFDocumento7 páginasMetabolismo de Lipídeos em Ruminantes PDFKaio FernandesAinda não há avaliações
- Atenção Nutricional Na Cirurgia Bariátrica e MetabólicaNo EverandAtenção Nutricional Na Cirurgia Bariátrica e MetabólicaNota: 5 de 5 estrelas5/5 (1)
- Soluções para Diabetes e Hipoglicemia (Traduzido): Como preveni-lo e livrar-se dele naturalmente, sem medicamentos, mas adotando um estilo de vida saudávelNo EverandSoluções para Diabetes e Hipoglicemia (Traduzido): Como preveni-lo e livrar-se dele naturalmente, sem medicamentos, mas adotando um estilo de vida saudávelAinda não há avaliações
- TCC Vanessa de Souza SteinDocumento38 páginasTCC Vanessa de Souza SteinAiran SilvaAinda não há avaliações
- 44 Alimentos e Bebidas AntienvelhecimentoDocumento39 páginas44 Alimentos e Bebidas AntienvelhecimentoMarcos Paulo Grillo100% (1)
- BIOQUIMICA - AULA 9 - Integração MetabólicaDocumento21 páginasBIOQUIMICA - AULA 9 - Integração MetabólicaLP BarbosaAinda não há avaliações
- Ingestão Moderada de Alimentos, Diabetes, Hipertensão e TabagismoDocumento30 páginasIngestão Moderada de Alimentos, Diabetes, Hipertensão e TabagismoJaceline BarrosAinda não há avaliações
- Diabetes - Quais Os Sintomas, Causas e Como Tratar185022Documento16 páginasDiabetes - Quais Os Sintomas, Causas e Como Tratar185022Fernanda pedro silvestre SilvestreAinda não há avaliações
- Empurrão Biologia 1Documento3 páginasEmpurrão Biologia 1profrenatobritoBastosNetoAinda não há avaliações
- Diabetes Mellitus SlideDocumento18 páginasDiabetes Mellitus SlideWemerson Sousa LimaAinda não há avaliações
- Gordura Corporal Total Conteúdo e Distribuição - En.ptDocumento25 páginasGordura Corporal Total Conteúdo e Distribuição - En.ptYara ZaninAinda não há avaliações
- Diabetes Mellitus TICS AfyaDocumento3 páginasDiabetes Mellitus TICS AfyaAndré MauricioAinda não há avaliações
- CAD e CHH 2022Documento14 páginasCAD e CHH 2022Jonas FerrazAinda não há avaliações
- Diabetes MellitusDocumento41 páginasDiabetes MellitusOrli Flávio PaternoAinda não há avaliações
- Apostila de DiabetesDocumento61 páginasApostila de DiabetesAmaury TrindadeAinda não há avaliações
- Alzheimer Diabetes Do Tipo 3Documento25 páginasAlzheimer Diabetes Do Tipo 3jessicaAinda não há avaliações
- Rev Port Diabetes 2018 - 13 - 49Documento13 páginasRev Port Diabetes 2018 - 13 - 49Nelimária Batista MoreiraAinda não há avaliações
- Diabete e MergulhoDocumento12 páginasDiabete e MergulhoRicardo GusmãoAinda não há avaliações
- Unidade 4 Herança MultifatorialDocumento29 páginasUnidade 4 Herança MultifatorialDiego AlarconAinda não há avaliações
- XigDuo Bula PacienteDocumento17 páginasXigDuo Bula PacienteegsavioAinda não há avaliações
- Receitas LowcarbDocumento15 páginasReceitas LowcarbRed SantosAinda não há avaliações
- Viva Com Saúde - Segunda EdiçãoDocumento61 páginasViva Com Saúde - Segunda EdiçãoGraciele FreitasAinda não há avaliações
- Guia Confidencial "O Fim Da Pressão Alta" Jolivi - Saúde NaturalDocumento18 páginasGuia Confidencial "O Fim Da Pressão Alta" Jolivi - Saúde NaturalFlavio AlvesAinda não há avaliações
- 11 Edição Emagrecimento Janeiro-2021Documento58 páginas11 Edição Emagrecimento Janeiro-2021alessandroAinda não há avaliações
- Artigo Carnes Vermelhas e CancerDocumento23 páginasArtigo Carnes Vermelhas e CancerGlauciaAinda não há avaliações
- Sobre Dantian - Efeitos Do Qigong Na Hemoglobina Glicada em Pacientes Com Diabetes Tipo 2. Um Estudo Prospetivo Randomizado Controlado Com PlaceboDocumento110 páginasSobre Dantian - Efeitos Do Qigong Na Hemoglobina Glicada em Pacientes Com Diabetes Tipo 2. Um Estudo Prospetivo Randomizado Controlado Com PlaceboBalbino SbAinda não há avaliações
- Diabetes ControladaDocumento105 páginasDiabetes Controladaandreoticas100% (3)
- Ebook GlicemiaDocumento23 páginasEbook GlicemiaRenato Oliveira VillarAinda não há avaliações
- Saúde Renovada - Aptidão FísicaDocumento4 páginasSaúde Renovada - Aptidão FísicaKaua MullerAinda não há avaliações
- Aula 4 - DiabetesDocumento8 páginasAula 4 - DiabetesEduardo KielingAinda não há avaliações
- 4 Passos para Controlar A DiabetesDocumento102 páginas4 Passos para Controlar A DiabetesControle da Diabetes BrasilAinda não há avaliações
- Aula DIABETES - Parte 2Documento67 páginasAula DIABETES - Parte 2pabline gerardi miguelAinda não há avaliações
- DIRETRIZ BRASILEIRA SOBRE DIABETES SlidesDocumento16 páginasDIRETRIZ BRASILEIRA SOBRE DIABETES SlidesNaiana RibeiroAinda não há avaliações