Você também pode gostar
- Sebenta TCMDocumento48 páginasSebenta TCMAnonymous mEU3GDIQDAinda não há avaliações
- O papel da proteína HIF-1α no desenvolvimento de tumoresDocumento40 páginasO papel da proteína HIF-1α no desenvolvimento de tumoresERGERRERG100% (1)
- FlipbookDocumento21 páginasFlipbookJohn JohnAinda não há avaliações
- Guia de Estudos Da Unidade 2 - Citologia e EmbriologiaDocumento13 páginasGuia de Estudos Da Unidade 2 - Citologia e EmbriologiaPastoral Familiar Nossa Senhora do CarmoAinda não há avaliações
- Estrutura da membrana21Documento6 páginasEstrutura da membrana21Monica AlexandreAinda não há avaliações
- Trabalaho Membrana PlasmáticaDocumento16 páginasTrabalaho Membrana PlasmáticaRonny GoisAinda não há avaliações
- Biologia Celular IntroduçãoDocumento38 páginasBiologia Celular IntroduçãoMaxwell M. RibeiroAinda não há avaliações
- AULA 3 BIOLOGIA CELULAR - Membranas Biologicas-1Documento81 páginasAULA 3 BIOLOGIA CELULAR - Membranas Biologicas-1Levi CamoesAinda não há avaliações
- Estrutura das Membranas CelularesDocumento19 páginasEstrutura das Membranas CelularesThiago SantosAinda não há avaliações
- Membrana Plasmática - Características e Resumo - Mundo EducaçãoDocumento3 páginasMembrana Plasmática - Características e Resumo - Mundo EducaçãoMarcos Vinícius Neves OlsanAinda não há avaliações
- Membrana CelularDocumento3 páginasMembrana CelularRodrigo KerberAinda não há avaliações
- PlementarDocumento3 páginasPlementarelizangelacarAinda não há avaliações
- A Membrana Plasmática É Formada BasicamenteDocumento5 páginasA Membrana Plasmática É Formada Basicamentearukalencar100% (6)
- Unid 1Documento23 páginasUnid 1renato bryanAinda não há avaliações
- Conceitos Gerais das Membranas CelularesDocumento8 páginasConceitos Gerais das Membranas CelularesDaniela GondimAinda não há avaliações
- Material de Apoio - Membrana Plasmática-Movimentação de Substâncias No Ambiente CelularDocumento14 páginasMaterial de Apoio - Membrana Plasmática-Movimentação de Substâncias No Ambiente CelularcitrolidaniAinda não há avaliações
- Organelas Celulares e Estruturas Celulares.Documento21 páginasOrganelas Celulares e Estruturas Celulares.Daiane LorenzonAinda não há avaliações
- Membrana celular: estrutura, composição e funçõesDocumento15 páginasMembrana celular: estrutura, composição e funçõesMARIA YASMIN DE SOUSA BERNARDOAinda não há avaliações
- Estrutura e funções da membrana celularDocumento10 páginasEstrutura e funções da membrana celularJulia AraujoAinda não há avaliações
- Componentes celularesDocumento17 páginasComponentes celularesVera DominguesAinda não há avaliações
- Membrana Plasmática. Digestão IntracelularDocumento27 páginasMembrana Plasmática. Digestão IntracelularBruno PereiraAinda não há avaliações
- Educação FísicaDocumento9 páginasEducação FísicaFatima aparecida de laraAinda não há avaliações
- Células eucariontes: estrutura e funçãoDocumento5 páginasCélulas eucariontes: estrutura e funçãoLucas NevesAinda não há avaliações
- Aula 4 - Biomembranas e OrganelasDocumento56 páginasAula 4 - Biomembranas e OrganelasxisbrjamillyAinda não há avaliações
- Estrutura e organelas da célula vegetalDocumento9 páginasEstrutura e organelas da célula vegetalCrisSumaqueiroAinda não há avaliações
- Transporte de substâncias nas célulasDocumento18 páginasTransporte de substâncias nas célulasMárcia JeteioAinda não há avaliações
- Resumo - GenéticaDocumento16 páginasResumo - Genéticaisabeljulia968Ainda não há avaliações
- Fisiologia VegetalDocumento8 páginasFisiologia VegetalRafael GuardadoAinda não há avaliações
- Curso Educação Física: Células e TecidosDocumento7 páginasCurso Educação Física: Células e TecidosNathan Toaldo SaporitoAinda não há avaliações
- A estrutura e função da membrana plasmáticaDocumento2 páginasA estrutura e função da membrana plasmáticaMaria Theresa MoreiraAinda não há avaliações
- Ebook Enem e Medicina Semana 3 Ao VivoDocumento419 páginasEbook Enem e Medicina Semana 3 Ao VivoNycolly NunesAinda não há avaliações
- Bases Biológicas Celulares e TecidaisDocumento14 páginasBases Biológicas Celulares e TecidaisMaria MariaAinda não há avaliações
- Estrutura e Funções da Membrana CelularDocumento3 páginasEstrutura e Funções da Membrana CelularMariana Gomes da Silva100% (1)
- Células eucariontes: estrutura e componentes da membrana plasmáticaDocumento18 páginasCélulas eucariontes: estrutura e componentes da membrana plasmáticaLucas NevesAinda não há avaliações
- Aula 4Documento12 páginasAula 4Aniceto RosarioAinda não há avaliações
- Material CITOLOGIA ORGANELASDocumento10 páginasMaterial CITOLOGIA ORGANELASScaryvanAinda não há avaliações
- Membrana Plasmá-WPS Office4Documento2 páginasMembrana Plasmá-WPS Office4Manuel KumissaAinda não há avaliações
- Instituto Politécnico PrivadoDocumento12 páginasInstituto Politécnico Privadosuedy pedroAinda não há avaliações
- Yan Mesko Da Silva - Atividade 6 - 1ºEM AnoDocumento3 páginasYan Mesko Da Silva - Atividade 6 - 1ºEM AnoYan Mesko05Ainda não há avaliações
- Atividade 3Documento7 páginasAtividade 3Priscila VidalAinda não há avaliações
- Comunicação e transporte celularDocumento33 páginasComunicação e transporte celularsem limitesAinda não há avaliações
- Membrana celular permeabilidadeDocumento21 páginasMembrana celular permeabilidadeAriadne SiqueiraAinda não há avaliações
- Fisiologia Das Membranas FMUPDocumento22 páginasFisiologia Das Membranas FMUPSónia RibeiroAinda não há avaliações
- Estudo Dirigido 2 - CitologiaDocumento4 páginasEstudo Dirigido 2 - CitologiaYara KerberAinda não há avaliações
- Membrana Celular - Composição e FunçõesDocumento9 páginasMembrana Celular - Composição e FunçõesElder Andrade AmaralAinda não há avaliações
- Membrana Celular: Estrutura, Funções e TransporteDocumento6 páginasMembrana Celular: Estrutura, Funções e Transporteta201tataAinda não há avaliações
- Membrana e organelas: estrutura e funçõesDocumento36 páginasMembrana e organelas: estrutura e funçõesAnna IzabelAinda não há avaliações
- Membrana celular: funções e estruturaDocumento20 páginasMembrana celular: funções e estruturaMario Maganizo JuniorAinda não há avaliações
- Modulo 2 Introducao A Citologia e Membrana Plasmatica - Doc - 20240314 - 113055 - 0000Documento13 páginasModulo 2 Introducao A Citologia e Membrana Plasmatica - Doc - 20240314 - 113055 - 0000Akiane FelixAinda não há avaliações
- Célula e suas estruturasDocumento5 páginasCélula e suas estruturasEdivaldoJrAinda não há avaliações
- Caio Maximus - Primeira Avaliação Do Curso de Biofísica Celular e Dos SistemasDocumento4 páginasCaio Maximus - Primeira Avaliação Do Curso de Biofísica Celular e Dos SistemasCaio MaximusAinda não há avaliações
- BIOMEMBRANASDocumento4 páginasBIOMEMBRANASaninhayomeAinda não há avaliações
- Membrana CelularDocumento5 páginasMembrana CelularMarcelo ColodetiAinda não há avaliações
- Turmadefevereiro-biologia1-Tipos de Células e Membrana Plasmática-12!04!2023-Fb6326a8fa586694572784406618a143Documento17 páginasTurmadefevereiro-biologia1-Tipos de Células e Membrana Plasmática-12!04!2023-Fb6326a8fa586694572784406618a143bio-luysasantosAinda não há avaliações
- Sistema MembranarDocumento7 páginasSistema MembranarAlteza Pró100% (3)
- AULA 4 BIOLOGIA CELULAR Estrutura Celular Membrana Plasmática Citoplasma NúcleoDocumento41 páginasAULA 4 BIOLOGIA CELULAR Estrutura Celular Membrana Plasmática Citoplasma Núcleocaroldsll14Ainda não há avaliações
- Biofísica das MembranasDocumento23 páginasBiofísica das MembranasWagner PimentelAinda não há avaliações
- CURSO DE EDUCAÇÃO FÍSICADocumento6 páginasCURSO DE EDUCAÇÃO FÍSICAGrazi MoreiraAinda não há avaliações
- Proteínas integrais da membrana plasmáticaDocumento3 páginasProteínas integrais da membrana plasmáticaJorge SalinasAinda não há avaliações
- Ed IDocumento4 páginasEd IMila MüllerAinda não há avaliações
- Membrana Plasmatica BioquimDocumento83 páginasMembrana Plasmatica Bioquimflavia douraoAinda não há avaliações
- Estresse Térmico SalinoDocumento15 páginasEstresse Térmico SalinoVivian CastilhoAinda não há avaliações
- Complexo de Golgi na Biologia CelularDocumento14 páginasComplexo de Golgi na Biologia CelularJuliana LeiteAinda não há avaliações
- BG10 - Teste Nº2A - 21nov2016 - CCDocumento3 páginasBG10 - Teste Nº2A - 21nov2016 - CCJogosAinda não há avaliações
- Relatório de aulas práticas sobre bioenergética, metabolismo e biologia celularDocumento5 páginasRelatório de aulas práticas sobre bioenergética, metabolismo e biologia celularDiego HenriqueAinda não há avaliações
- Revisão ClostridosesDocumento20 páginasRevisão ClostridosesAmanda HervisAinda não há avaliações
- BiofotossínteseDocumento21 páginasBiofotossínteseGuilherme Souza GonçalvesAinda não há avaliações
- Livro - Biologia, Histologia e EmbriologiaDocumento52 páginasLivro - Biologia, Histologia e EmbriologiaJoão PedroAinda não há avaliações
- Exercícios sobre tecido muscular e organelas 1o ano ensino médioDocumento17 páginasExercícios sobre tecido muscular e organelas 1o ano ensino médioThiago CostaAinda não há avaliações
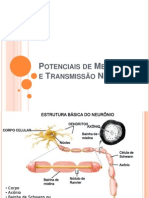
- 6.potenciais de Membrana e Transmissao NervosaDocumento30 páginas6.potenciais de Membrana e Transmissao NervosaDenny Vitor Barbosa RamosAinda não há avaliações
- Estudo Dirigido de Aulas Práticas - Fag ParanáDocumento31 páginasEstudo Dirigido de Aulas Práticas - Fag ParanáVanessaDoEsa100% (1)
- Anatomia VeterináriaDocumento103 páginasAnatomia VeterináriaFrances FreitasAinda não há avaliações
- Robbins & Cotran Fundamentos de PatologiaDocumento1.201 páginasRobbins & Cotran Fundamentos de PatologiaGiovana RodriguesAinda não há avaliações
- Biologia celular e citologia clínicaDocumento37 páginasBiologia celular e citologia clínicaLidiane PenaAinda não há avaliações
- Portfolio LucasDocumento13 páginasPortfolio LucaslucasAinda não há avaliações
- Livroatlasbiocel PDFDocumento138 páginasLivroatlasbiocel PDFigorAinda não há avaliações
- Diferenças Entre Célula Animal e VegetalDocumento1 páginaDiferenças Entre Célula Animal e VegetalDavi RafaelAinda não há avaliações
- Bio Campbell-1-500-151-200Documento50 páginasBio Campbell-1-500-151-200AsukaAinda não há avaliações
- Citologia 2013 Citoplasma 2013 Sintese Armazenamento e TransporteDocumento27 páginasCitologia 2013 Citoplasma 2013 Sintese Armazenamento e TransporteBeboc Deagonal UlfricoAinda não há avaliações
- Monensina e virginiamicina para bovinos de corte: desempenho e simulação econômicaDocumento25 páginasMonensina e virginiamicina para bovinos de corte: desempenho e simulação econômicaPedro SantosAinda não há avaliações
- Estrutura e componentes da célula eucariotaDocumento3 páginasEstrutura e componentes da célula eucariotaRogério CanoasAinda não há avaliações
- Anatomia Funcional Das Células BacterianasDocumento40 páginasAnatomia Funcional Das Células Bacterianasviniciusbarbosa8090Ainda não há avaliações
- Farmacocinética e FarmacodinâmicaDocumento3 páginasFarmacocinética e FarmacodinâmicaLaís Almeida100% (1)
- Organelas celulares: uma viagem pelo mundo intracelularDocumento5 páginasOrganelas celulares: uma viagem pelo mundo intracelularAshleyAinda não há avaliações
- Membranas biológicas: estrutura e funções emDocumento3 páginasMembranas biológicas: estrutura e funções emGabriel OliveiraAinda não há avaliações
- Toxicologia UNIPDocumento45 páginasToxicologia UNIPJonas CostaAinda não há avaliações
- Prova - Bioquímica - UFABCDocumento2 páginasProva - Bioquímica - UFABCFelipe OliveiraAinda não há avaliações
- 4º Relatório de Biologia CelularDocumento11 páginas4º Relatório de Biologia CelularalinemsantanaAinda não há avaliações
- C1 CursoE Biologia 20aulasDocumento24 páginasC1 CursoE Biologia 20aulasHancock8683% (6)