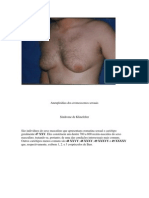
Você também pode gostar
- Gene SRY e determinação sexualDocumento9 páginasGene SRY e determinação sexualJosefa MoreiraAinda não há avaliações
- Análise Do ArtigoDocumento15 páginasAnálise Do ArtigoAna MartinsAinda não há avaliações
- Diferenciação Entre Os Cérebros Masculino e FemininoDocumento5 páginasDiferenciação Entre Os Cérebros Masculino e FemininoNara NeylaineAinda não há avaliações
- Cromossoma Y e o Fator Determinante Do TestículoDocumento9 páginasCromossoma Y e o Fator Determinante Do TestículoCássio QueirozAinda não há avaliações
- Diferenciação do cérebro masculino e femininoDocumento4 páginasDiferenciação do cérebro masculino e femininoJosidarkAinda não há avaliações
- Gônadas de Indivíduos Com Distúrbio Do Desenvolvimento Sexual Ovotesticular: Uma Revisão SistemáticaDocumento10 páginasGônadas de Indivíduos Com Distúrbio Do Desenvolvimento Sexual Ovotesticular: Uma Revisão SistemáticaFranklin de OliveiraAinda não há avaliações
- Aula 4 Comportamento ReprodutivoDocumento27 páginasAula 4 Comportamento ReprodutivoAngelica Edson OliveiraAinda não há avaliações
- Principais descobertas sobre infertilidade masculinaDocumento3 páginasPrincipais descobertas sobre infertilidade masculinacatarina rodriguesAinda não há avaliações
- Gene SRYDocumento1 páginaGene SRYDebora ReisAinda não há avaliações
- TICs DIFERNCIAÇÃO GONADALDocumento3 páginasTICs DIFERNCIAÇÃO GONADALMaria Vitória DominguesAinda não há avaliações
- Pedofilia - Considerações atuais sobre etiologia e tratamentoDocumento2 páginasPedofilia - Considerações atuais sobre etiologia e tratamentoDanielle NunesAinda não há avaliações
- Determinação Sexual Aula 09.06 - Parte 2Documento2 páginasDeterminação Sexual Aula 09.06 - Parte 2Stefani CostaAinda não há avaliações
- O que é Hermafroditismo? Entendendo as causas e tratamentos do intersexoDocumento4 páginasO que é Hermafroditismo? Entendendo as causas e tratamentos do intersexoBassalo2018Ainda não há avaliações
- Diferenciação sexual normal e anômalaDocumento3 páginasDiferenciação sexual normal e anômalaDescomplica ENEMAinda não há avaliações
- Genitália AmbiguaDocumento11 páginasGenitália AmbiguaTauan de OliveiraAinda não há avaliações
- As bases biológicas das diferenças entre sexosDocumento24 páginasAs bases biológicas das diferenças entre sexosCatuta Azarado Paizinho ErnestoAinda não há avaliações
- BiologiaDocumento22 páginasBiologiaRômulo FernandesAinda não há avaliações
- Susanesb,+Revista SBRASH ECDocumento7 páginasSusanesb,+Revista SBRASH ECBianca Quattrer Alves ribeiroAinda não há avaliações
- Síndrome de Klinefelter: Causas, Sinais e DiagnósticoDocumento11 páginasSíndrome de Klinefelter: Causas, Sinais e DiagnósticoLarissa OliveiraAinda não há avaliações
- Nal 1Documento12 páginasNal 1Diacuires Amaral DantasAinda não há avaliações
- Teste de Avaliacao Escrita de Biologia 12 1 ReservaDocumento7 páginasTeste de Avaliacao Escrita de Biologia 12 1 ReservaRenato Costa100% (21)
- Sexologia forense: estudo dos fenômenos relacionados à reprodução humanaDocumento22 páginasSexologia forense: estudo dos fenômenos relacionados à reprodução humanath1ag0rsAinda não há avaliações
- Diferença SexualDocumento2 páginasDiferença SexualFlávio DavidAinda não há avaliações
- Determinação sexual e diferenciação gonadal no embrião e fetoDocumento10 páginasDeterminação sexual e diferenciação gonadal no embrião e fetomateus_z100% (1)
- Diferenciação Biológica Do Sexo e Do Sistema GenitalDocumento39 páginasDiferenciação Biológica Do Sexo e Do Sistema GenitalFelipe BastosAinda não há avaliações
- Diferença Entre Os Sexos Na EsquizofreniaDocumento2 páginasDiferença Entre Os Sexos Na EsquizofreniaLaura CostaAinda não há avaliações
- O ponto X do sexo: seqüenciamento do cromossomo X humanoDocumento2 páginasO ponto X do sexo: seqüenciamento do cromossomo X humanodsscecilioAinda não há avaliações
- Infertilidade masculina: causas e descobertasDocumento15 páginasInfertilidade masculina: causas e descobertascatarina rodriguesAinda não há avaliações
- Determinação Genética do Sexo na Espécie HumanaDocumento13 páginasDeterminação Genética do Sexo na Espécie HumanaGusta MonteiroAinda não há avaliações
- Homem XX Relato de Três Casos Na FaixaDocumento4 páginasHomem XX Relato de Três Casos Na FaixaJulliana Leocádio Bastos RangelAinda não há avaliações
- 10093-Texto Do Artigo-54806-2-10-20211104Documento13 páginas10093-Texto Do Artigo-54806-2-10-20211104thsoffnerAinda não há avaliações
- Aspectos moleculares da determinação e diferenciação sexualDocumento11 páginasAspectos moleculares da determinação e diferenciação sexualPatrícia NettoAinda não há avaliações
- 12 - Autismo, TDAH, Vício e Diferenciação Sexual Do Sistema NervosoDocumento18 páginas12 - Autismo, TDAH, Vício e Diferenciação Sexual Do Sistema NervosoeduardosmourasAinda não há avaliações
- Síndrome de Klinefelter: aspectos genotípicos, fenotípicos e dimensões psicossociaisDocumento7 páginasSíndrome de Klinefelter: aspectos genotípicos, fenotípicos e dimensões psicossociaisBárbaraRafaelaAinda não há avaliações
- Apostila I - O Eneagrama PDFDocumento68 páginasApostila I - O Eneagrama PDFMiryan C. Chagas100% (2)
- Endocrinologia Masturbação e Homossexualidade 1 PDocumento38 páginasEndocrinologia Masturbação e Homossexualidade 1 PsandraAinda não há avaliações
- Cariótipo: Análise de CromossomosDocumento3 páginasCariótipo: Análise de CromossomosJuan LucasAinda não há avaliações
- Resumo Do Livro O Cromossomo Y e A Biologia Das Celulas Germinativas Masculinas Na Saude e Nas Doenças - 2007Documento42 páginasResumo Do Livro O Cromossomo Y e A Biologia Das Celulas Germinativas Masculinas Na Saude e Nas Doenças - 2007SELMA DOS SANTOS CAROLINO DE ANDRADEAinda não há avaliações
- Citogenética ClínicaDocumento30 páginasCitogenética ClínicaMaria Clara FernandesAinda não há avaliações
- A Epigenetica e A Transmissão PsiquicaDocumento15 páginasA Epigenetica e A Transmissão Psiquicanadiabeatriz.qAinda não há avaliações
- Síndrome KlinefelterDocumento6 páginasSíndrome KlinefelterGabrielle GonçalvesAinda não há avaliações
- Anomalias CromossômicasDocumento9 páginasAnomalias CromossômicasMARCELO FERREIRA100% (2)
- Estágio de Citogenética2Documento56 páginasEstágio de Citogenética2Fenias BoaneAinda não há avaliações
- Sexagem de sêmen: biotecnologia para direcionar o sexo dos bezerrosDocumento11 páginasSexagem de sêmen: biotecnologia para direcionar o sexo dos bezerrosHannah CecíliaAinda não há avaliações
- DiferSEX anomaliasDocumento2 páginasDiferSEX anomaliasLuis Eduardo M Faria100% (1)
- Hermafroditismo Verdadeiro: 36 CasosDocumento8 páginasHermafroditismo Verdadeiro: 36 Casosluanalima2000Ainda não há avaliações
- Hermafroditismo: causas, tipos e tratamentoDocumento1 páginaHermafroditismo: causas, tipos e tratamentoNo OneAinda não há avaliações
- 1 Desen Embrio e Difer SexualDocumento17 páginas1 Desen Embrio e Difer SexualChácara Arte SuaveAinda não há avaliações
- Eterminação Romossômica Do EXO: Prof . Ms. Claudia GiaquintoDocumento37 páginasEterminação Romossômica Do EXO: Prof . Ms. Claudia GiaquintoninjagamereytAinda não há avaliações
- Artigo - Jose de Lima Machado - 2013Documento4 páginasArtigo - Jose de Lima Machado - 2013Marcos Gonçalves AlmeidaAinda não há avaliações
- Porque o Y É Tão EstranhoDocumento2 páginasPorque o Y É Tão EstranhoRodolfo Antônio da SilvaAinda não há avaliações
- Normas para tratamento de anomalias de diferenciação sexualDocumento5 páginasNormas para tratamento de anomalias de diferenciação sexualDeise BrasilAinda não há avaliações
- Aula 8 - Determinação Do Sexo Na Espécie HumanaDocumento16 páginasAula 8 - Determinação Do Sexo Na Espécie HumanaEVERTONAinda não há avaliações
- Considerações Psicanalíticas Sobre A Mulher' Na C - 230427 - 133128Documento23 páginasConsiderações Psicanalíticas Sobre A Mulher' Na C - 230427 - 133128RaiannenAinda não há avaliações
- O BIG BANG DA VIDA - o Estudo Biopsicossocial Da Concepção Ao NascimentoDocumento18 páginasO BIG BANG DA VIDA - o Estudo Biopsicossocial Da Concepção Ao NascimentoPatriciaAinda não há avaliações
- A Importância Dos Aspectos Éticos No IntersexoDocumento14 páginasA Importância Dos Aspectos Éticos No IntersexoJuliana AlvesAinda não há avaliações
- Teste de Biologia sobre Espermatogénese e Ciclo Sexual FemininoDocumento7 páginasTeste de Biologia sobre Espermatogénese e Ciclo Sexual FemininostepoutnowAinda não há avaliações
- apostila1-machoDocumento56 páginasapostila1-machodanielleferreirat2Ainda não há avaliações
- 7 perguntas ainda não respondidas pela FísicaDocumento13 páginas7 perguntas ainda não respondidas pela FísicaisabutterflyloverAinda não há avaliações
- 5 traços que identificam um agressor psicológicoDocumento11 páginas5 traços que identificam um agressor psicológicoisabutterflyloverAinda não há avaliações
- Uma mulher inteligente sabe que não tem limitesDocumento12 páginasUma mulher inteligente sabe que não tem limitesisabutterflyloverAinda não há avaliações
- Síndrome de ilegitimidadeDocumento12 páginasSíndrome de ilegitimidadeisabutterflyloverAinda não há avaliações
- A fascinante história do DSMDocumento11 páginasA fascinante história do DSMisabutterflyloverAinda não há avaliações
- 6 formas de sabotagem do egoDocumento11 páginas6 formas de sabotagem do egoisabutterflyloverAinda não há avaliações
- Você não é o que você conquistaDocumento13 páginasVocê não é o que você conquistaisabutterflyloverAinda não há avaliações
- 3 estratégias para aumentar a autoestimaDocumento11 páginas3 estratégias para aumentar a autoestimaisabutterflyloverAinda não há avaliações
- Karl Marx e A Diferença Entre Comunismo e SocialismoDocumento7 páginasKarl Marx e A Diferença Entre Comunismo e SocialismoisabutterflyloverAinda não há avaliações
- AmbientalismoDocumento15 páginasAmbientalismoisabutterflyloverAinda não há avaliações
- NataçãoDocumento12 páginasNataçãoisabutterflyloverAinda não há avaliações
- O Mundo PósDocumento7 páginasO Mundo PósisabutterflyloverAinda não há avaliações
- Comuna de ParisDocumento4 páginasComuna de ParisisabutterflyloverAinda não há avaliações
- Estudo 01 - Que É Batismo Com o Espirito SantoDocumento3 páginasEstudo 01 - Que É Batismo Com o Espirito SantoHenrique JorgeAinda não há avaliações
- Tutorial LPIDocumento3 páginasTutorial LPIrandomerdreamerAinda não há avaliações
- Primeiros Socorros Básico ProvaDocumento5 páginasPrimeiros Socorros Básico ProvaDcastro Soares80% (5)
- Normas para apresentação de ficheiros digitais em processos urbanísticosDocumento18 páginasNormas para apresentação de ficheiros digitais em processos urbanísticosPicaPauAmareloAinda não há avaliações
- aÇÃo de Pedido de RemÉdioDocumento15 páginasaÇÃo de Pedido de RemÉdioapi-3798293100% (9)
- Mais Que AmigosDocumento1 páginaMais Que AmigosRafael SathlerAinda não há avaliações
- CitologiaDocumento43 páginasCitologiawolfgangvieira100% (1)
- Equipamentos para corte e recolha de forragensDocumento26 páginasEquipamentos para corte e recolha de forragensAndré QuirinoAinda não há avaliações
- Aula 09.replicação Do DNADocumento42 páginasAula 09.replicação Do DNArmdsAinda não há avaliações
- Regras lotação servidores educação AMDocumento6 páginasRegras lotação servidores educação AMDanielaCarvalhoAinda não há avaliações
- Radiestesia - Reiki - e - Radiestesia - ApostilaDocumento368 páginasRadiestesia - Reiki - e - Radiestesia - ApostilaANDRÔMEDA.Ainda não há avaliações
- EMH FREIO A DISCO MANUAL MANUTENÇÃO FDE COM ALAVANCA Rev SensorDocumento29 páginasEMH FREIO A DISCO MANUAL MANUTENÇÃO FDE COM ALAVANCA Rev SensorArnaldo Mazzei100% (3)
- RESENHA em Busca Da Mente MusicalDocumento8 páginasRESENHA em Busca Da Mente MusicalKátia GallyAinda não há avaliações
- EJA e sua importância socialDocumento82 páginasEJA e sua importância socialJose Roberto Junior Costa100% (1)
- MovTer-cortesaterrostaludesDocumento51 páginasMovTer-cortesaterrostaludesdouglasAinda não há avaliações
- Ótica UFSC lista exercícios refração luzDocumento3 páginasÓtica UFSC lista exercícios refração luzRommel AscenciónAinda não há avaliações
- Plano de Atividade de Estágio Não ObrigatórioDocumento3 páginasPlano de Atividade de Estágio Não ObrigatórioJose CiceroAinda não há avaliações
- Texto 04 - QuadriláterosDocumento8 páginasTexto 04 - QuadriláterosAbílio Júnior MubaiAinda não há avaliações
- 1946 - 1964 A Experiência Democrática No BrasilDocumento8 páginas1946 - 1964 A Experiência Democrática No BrasilJosé Renato TeixeiraAinda não há avaliações
- Aula de Desfibrilador-Cardioversor EC TranspDocumento15 páginasAula de Desfibrilador-Cardioversor EC Transpjosuemax3Ainda não há avaliações
- Doc3 - ISO 9001 - Comunicação Interna PDFDocumento3 páginasDoc3 - ISO 9001 - Comunicação Interna PDFzeliaornelasAinda não há avaliações
- Roteiro Aula Pratica P EnviarDocumento4 páginasRoteiro Aula Pratica P EnviarÉrika PatriciaAinda não há avaliações
- Avaliação intercalar EB 2,3 VisoDocumento1 páginaAvaliação intercalar EB 2,3 VisoprofalexramosAinda não há avaliações
- Uma Família Firmada Na Rocha Parte FinalDocumento2 páginasUma Família Firmada Na Rocha Parte FinalREINÃ CASTILHO MARTINSAinda não há avaliações
- O Poder Das Ervas Na África - Selma Sá e Rodrigo de Logun - Versão Final - CodDocumento85 páginasO Poder Das Ervas Na África - Selma Sá e Rodrigo de Logun - Versão Final - CodDaniel R.O100% (8)
- Prefeitura Municipal de Vila-Velha Estado Do Espiríto Santo Secretaria Municipal de EducaçãoDocumento4 páginasPrefeitura Municipal de Vila-Velha Estado Do Espiríto Santo Secretaria Municipal de EducaçãoErica Dos Santos MascarenhasAinda não há avaliações
- Aposto - VocativoDocumento11 páginasAposto - VocativoAlessandra CoutoAinda não há avaliações
- Heterósidos cardíacos: estrutura, classificação e atividadeDocumento30 páginasHeterósidos cardíacos: estrutura, classificação e atividadeIsabela Honorato100% (1)
- 6ºano Slides Aula 03Documento12 páginas6ºano Slides Aula 03MICHIELI OLIVEIRA RIBEIROAinda não há avaliações
- Biomecânica Básica dos Exercícios: Membros InferioresNo EverandBiomecânica Básica dos Exercícios: Membros InferioresNota: 3.5 de 5 estrelas3.5/5 (8)
- Hackeando Darwin: Engenharia genética e o futuro da humanidadeNo EverandHackeando Darwin: Engenharia genética e o futuro da humanidadeAinda não há avaliações
- Emoções: Introdução à psicologia das emoções. De Darwin à neurociência, o que são as emoções e como elas funcionamNo EverandEmoções: Introdução à psicologia das emoções. De Darwin à neurociência, o que são as emoções e como elas funcionamNota: 5 de 5 estrelas5/5 (4)
- E-TRAP: entrevista diagnóstica para transtornos de personalidadeNo EverandE-TRAP: entrevista diagnóstica para transtornos de personalidadeNota: 5 de 5 estrelas5/5 (3)
- Treinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNo EverandTreinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNota: 4.5 de 5 estrelas4.5/5 (169)
- Aprender mais rápido: Habilidades de estudo, memória e neurociênciaNo EverandAprender mais rápido: Habilidades de estudo, memória e neurociênciaNota: 4 de 5 estrelas4/5 (5)
- Psicoterapia: Teorias e técnicas psicoterápicasNo EverandPsicoterapia: Teorias e técnicas psicoterápicasNota: 5 de 5 estrelas5/5 (4)
- Uma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNo EverandUma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNota: 4 de 5 estrelas4/5 (13)
- Suplementação proteica: Aminoacidemia e saúde vascularNo EverandSuplementação proteica: Aminoacidemia e saúde vascularAinda não há avaliações
- Educação ambiental: A formação do sujeito ecológicoNo EverandEducação ambiental: A formação do sujeito ecológicoAinda não há avaliações
- O Licenciamento Ambiental no Brasil na PráticaNo EverandO Licenciamento Ambiental no Brasil na PráticaNota: 1 de 5 estrelas1/5 (2)
- Abismos Narcísicos: A Psicodinâmica do Amadurecimento e da IndividuaçãoNo EverandAbismos Narcísicos: A Psicodinâmica do Amadurecimento e da IndividuaçãoNota: 5 de 5 estrelas5/5 (2)
- Tudo tem uma explicação: A biologia por trás de tudo aquilo que você nunca imaginouNo EverandTudo tem uma explicação: A biologia por trás de tudo aquilo que você nunca imaginouNota: 4 de 5 estrelas4/5 (5)
- Reflexões sobre o Luto: Práticas Interventivas e Especificidades do Trabalho com Pessoas EnlutadasNo EverandReflexões sobre o Luto: Práticas Interventivas e Especificidades do Trabalho com Pessoas EnlutadasAinda não há avaliações