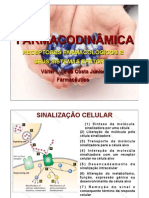
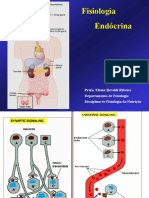
Você também pode gostar
- Prolactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoNo EverandProlactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoAinda não há avaliações
- Artigo - Vias de Sinalização Da InsulinaDocumento23 páginasArtigo - Vias de Sinalização Da InsulinaArthur PetronettoAinda não há avaliações
- SinalizaçãoDocumento4 páginasSinalizaçãoMariana PaesAinda não há avaliações
- Aula 5 - Receptores Farmacológicos e Sistemas EfetoresDocumento27 páginasAula 5 - Receptores Farmacológicos e Sistemas EfetoresRodolfo Rodrigues Zacarias100% (4)
- BVE 3009 - AULA 1-Conceitos de Crescimento e DesenvolvimentoDocumento44 páginasBVE 3009 - AULA 1-Conceitos de Crescimento e DesenvolvimentoLaís VictóriaAinda não há avaliações
- Aula 6 - Transdução de Sinais BiológicosDocumento12 páginasAula 6 - Transdução de Sinais BiológicosJailton MagalhãesAinda não há avaliações
- Resumo 4 Receptores e A Transdução de SinalDocumento8 páginasResumo 4 Receptores e A Transdução de Sinalthaisa bocattiAinda não há avaliações
- SinalizaçãoDocumento11 páginasSinalizaçãoIsmael SilveiraAinda não há avaliações
- Biossinalização - Beatriz MayumiDocumento3 páginasBiossinalização - Beatriz Mayumibe.mayumi.2010Ainda não há avaliações
- Atividades de BiossinalizaçãoDocumento3 páginasAtividades de Biossinalizaçãobe.mayumi.2010Ainda não há avaliações
- Vias de Sinalização Que Controlam A Expressão GênicaDocumento8 páginasVias de Sinalização Que Controlam A Expressão GênicaJoao ManoelAinda não há avaliações
- BiossinalizaçãoDocumento17 páginasBiossinalizaçãoMelissaAlvesAinda não há avaliações
- Síntese e Degradação de NeurotransmissoresDocumento75 páginasSíntese e Degradação de NeurotransmissoresEvanilson de LimaAinda não há avaliações
- Mecanismos Moleculares de Resistência À Insulina Na Síndrome MetabólicaDocumento23 páginasMecanismos Moleculares de Resistência À Insulina Na Síndrome MetabólicaSandra Carolina Salguero Cruz100% (1)
- Farmacologia Resumo ReceptologiaDocumento15 páginasFarmacologia Resumo ReceptologiaPedro MeirelesAinda não há avaliações
- Comunicacao e Sinalizacao Celular ApontamentosDocumento15 páginasComunicacao e Sinalizacao Celular Apontamentos708242287Ainda não há avaliações
- BQI 241 OkDocumento13 páginasBQI 241 OkCaio AugustusAinda não há avaliações
- Aula9 Expressao Genica EucariotosDocumento63 páginasAula9 Expressao Genica EucariotosCoordenação Ciências BiológicasAinda não há avaliações
- FARMACOLOGIA II 07 - Autacóides (Anti-Histamínicos) - MAIO-2011Documento6 páginasFARMACOLOGIA II 07 - Autacóides (Anti-Histamínicos) - MAIO-2011Beatriz GuimarãesAinda não há avaliações
- Análise de Micro RNAs No Hipotálamo de Camundongos C57BL/6 Treinados Alimentados Com Dieta HiperlipídicaDocumento16 páginasAnálise de Micro RNAs No Hipotálamo de Camundongos C57BL/6 Treinados Alimentados Com Dieta HiperlipídicaJessica SofiattiAinda não há avaliações
- Guião CBL 1Documento53 páginasGuião CBL 1Diana BatistaAinda não há avaliações
- Biologia Molecular e Celular - Resumo Da Prova 2 PDFDocumento23 páginasBiologia Molecular e Celular - Resumo Da Prova 2 PDFIvan boavidaAinda não há avaliações
- Mecanismo de Ação Dos Fármacos: 06/02/2023 FarmacologiaDocumento7 páginasMecanismo de Ação Dos Fármacos: 06/02/2023 FarmacologiaCamila Goncalves MirandaAinda não há avaliações
- Sinalização CelularDocumento6 páginasSinalização CelularChristianAinda não há avaliações
- Receptores Enzimáticos e Associados À EnzimasDocumento6 páginasReceptores Enzimáticos e Associados À EnzimasMax Nunes100% (1)
- Luis Augusto AInesDocumento44 páginasLuis Augusto AInesVictória PatríciaAinda não há avaliações
- Vias de Sinalização Da InsulinaDocumento5 páginasVias de Sinalização Da InsulinaNeide Dumas Figueiredo100% (1)
- Mecanismos de Transdução de Sinal para Os Receptores de Membrana PlasmáticaDocumento12 páginasMecanismos de Transdução de Sinal para Os Receptores de Membrana PlasmáticaJacqueline Quintal100% (1)
- 02 - Classificação Dos HormôniosDocumento3 páginas02 - Classificação Dos HormôniosLuis OliveiraAinda não há avaliações
- Farmacologia EndócrinaDocumento12 páginasFarmacologia EndócrinaLamguri Robertsoniano100% (1)
- Aula 8Documento23 páginasAula 8Diogo CostaAinda não há avaliações
- Fatores de Crescimento PDFDocumento5 páginasFatores de Crescimento PDFAlexandredeAguiar100% (1)
- HormonasDocumento31 páginasHormonasxicana83% (6)
- Integração MetabólicaDocumento35 páginasIntegração MetabólicaRicardo BorgesAinda não há avaliações
- Sinalização Celular No Cancer PDFDocumento4 páginasSinalização Celular No Cancer PDFJhonny BiologoAinda não há avaliações
- Receptores CelularesDocumento29 páginasReceptores CelularesMarina Giusepette100% (1)
- 01 2018 Penicilinas E Inibidores Da BetalactamaseDocumento123 páginas01 2018 Penicilinas E Inibidores Da BetalactamaseGabryel GomesAinda não há avaliações
- Tese - ProlactinaDocumento80 páginasTese - ProlactinamicalAinda não há avaliações
- Aula Mecanismo EndocDocumento52 páginasAula Mecanismo EndocabresserAinda não há avaliações
- Aula de HormôniosDocumento31 páginasAula de HormôniosDanielle A. M. MatteoAinda não há avaliações
- Endócrino e Nutrição Bianca RamalloDocumento219 páginasEndócrino e Nutrição Bianca RamalloVivian Tezzelle100% (2)
- ED 2 - Farmacologia Aplicada Ao Sistema ColinérgicoDocumento6 páginasED 2 - Farmacologia Aplicada Ao Sistema ColinérgicoAdele LeiteAinda não há avaliações
- Dicionário de Imunologia PDFDocumento361 páginasDicionário de Imunologia PDFLemuel DinizAinda não há avaliações
- Resumos Farmacologia 2 FrequênciaDocumento19 páginasResumos Farmacologia 2 FrequênciaBeatriz RebeloAinda não há avaliações
- Soares 2003Documento6 páginasSoares 2003ian souzaAinda não há avaliações
- 1.2 Mecanismo de Transdução HormonalDocumento3 páginas1.2 Mecanismo de Transdução HormonalJaqueline SantosAinda não há avaliações
- 2 Aula FarmadinamicaDocumento48 páginas2 Aula FarmadinamicaEduardo CostaAinda não há avaliações
- Aula 5 - Sinalização CelularDocumento37 páginasAula 5 - Sinalização CelularCamila HerculanoAinda não há avaliações
- BioquimicaDocumento90 páginasBioquimicaFlorAinda não há avaliações
- Proteina GDocumento6 páginasProteina GThalyta Pinheiro MedeirosAinda não há avaliações
- Proteínas Tirosinas Quinases - Desafios Do Desenvolvimento de Fármacos para A Terapia Do CâncerDocumento24 páginasProteínas Tirosinas Quinases - Desafios Do Desenvolvimento de Fármacos para A Terapia Do CâncerRenan Guilherme de Oliveira GuihAinda não há avaliações
- NutricaoDocumento80 páginasNutricaobruna vilelaAinda não há avaliações
- Tipos de Sinapses ModulatóriasDocumento5 páginasTipos de Sinapses ModulatóriasMrkinghtgAinda não há avaliações
- LeptinaDocumento17 páginasLeptinaCarlos EduardoAinda não há avaliações
- 04 - Introdução À Enzimologia e Dosagem de Enzimas Específicas No SoroDocumento27 páginas04 - Introdução À Enzimologia e Dosagem de Enzimas Específicas No SoroAndréia VieiraAinda não há avaliações
- 19.1 Sinalizaçao CelularDocumento6 páginas19.1 Sinalizaçao CelularAna CristinaAinda não há avaliações
- Resistência À Insulina No Diabetes Tipo 2Documento16 páginasResistência À Insulina No Diabetes Tipo 2Felipe BönmannAinda não há avaliações
- BiossinalizaçãoDocumento19 páginasBiossinalizaçãoSergio Correa FernandesAinda não há avaliações
- Controle de Expressão Gênica em EucariotosDocumento51 páginasControle de Expressão Gênica em EucariotosVittor GuidoniAinda não há avaliações
- Biomol - CitoesqueletoDocumento2 páginasBiomol - Citoesqueletocarol pimentelAinda não há avaliações
- Gliconeogênese Seminario Rodolfo 29 de Outubro 2021.2Documento7 páginasGliconeogênese Seminario Rodolfo 29 de Outubro 2021.2MC CHACALAinda não há avaliações
- Lâmina - CactinDocumento2 páginasLâmina - CactinCaiqueAinda não há avaliações
- Componentes Químicos Da CélulaDocumento30 páginasComponentes Químicos Da CélulasalariortegaAinda não há avaliações
- FSA - Aula - 01 - Vitamina e o CérebroDocumento72 páginasFSA - Aula - 01 - Vitamina e o CérebroLiliana CarvalhoAinda não há avaliações
- Sliedes Aula Carboidratos Não FibrososDocumento23 páginasSliedes Aula Carboidratos Não FibrososRodrigo RossiAinda não há avaliações
- BioquimicaDocumento6 páginasBioquimicaana290392Ainda não há avaliações
- 2 - Ficha de Exercicios.Documento4 páginas2 - Ficha de Exercicios.Adão Bila IdalécioAinda não há avaliações
- Jeferson 2022Documento9 páginasJeferson 2022Ronaldo Bastos100% (1)
- 5.1 Suplementação Atualizada Cosntipação Infantil + Suplementação 2 A 6 AnosDocumento2 páginas5.1 Suplementação Atualizada Cosntipação Infantil + Suplementação 2 A 6 AnosAlemao LeonHeartAinda não há avaliações
- 2-Questões BQ II - Vitaminas 2 PDFDocumento4 páginas2-Questões BQ II - Vitaminas 2 PDFBeatriz SPAinda não há avaliações
- 22 - 23 - 24-Slides-Ciclo - Alimentacao-Jejum-2014-4por PagDocumento15 páginas22 - 23 - 24-Slides-Ciclo - Alimentacao-Jejum-2014-4por PagJanaina Cunha Barbosa DalloAinda não há avaliações
- Livro Lehningher - ÍndiceDocumento1 páginaLivro Lehningher - ÍndiceIngrid CQAinda não há avaliações
- Questões Concurso Bromatologia 2º BimestreDocumento11 páginasQuestões Concurso Bromatologia 2º BimestreCibelle CarvalhoAinda não há avaliações
- Bioquimica Met TUDO JUNTODocumento44 páginasBioquimica Met TUDO JUNTOI Love DepilAinda não há avaliações
- ANTIDISLIPIDEMICOSDocumento10 páginasANTIDISLIPIDEMICOSLuiza StangerAinda não há avaliações
- 13 - LipogêneseDocumento4 páginas13 - Lipogênesemaria eduarda100% (1)
- Proteínas 2 2Documento57 páginasProteínas 2 2Isabela RochaAinda não há avaliações
- Funções Dos LipídeosDocumento3 páginasFunções Dos LipídeosGabriel MirandaAinda não há avaliações
- Esquema Das BoimolélulasDocumento2 páginasEsquema Das BoimolélulasGéssica GutierresAinda não há avaliações
- Novo ApresentaçãoDocumento11 páginasNovo ApresentaçãoandrechicoAinda não há avaliações
- DogmaresolDocumento6 páginasDogmaresolCaroline XavierAinda não há avaliações
- Ficha Tecnica Oleo de CanolaDocumento1 páginaFicha Tecnica Oleo de CanolaVstregaAinda não há avaliações
- Lista de Exercícios - Bioquímica (Lipídeos Ou Lipídios) : 01 - (UECE)Documento17 páginasLista de Exercícios - Bioquímica (Lipídeos Ou Lipídios) : 01 - (UECE)Kristiano RatzingerAinda não há avaliações
- Aula 02 - Aminoácidos e Proteínas-1Documento54 páginasAula 02 - Aminoácidos e Proteínas-1Gabriel Bahia dos AnjosAinda não há avaliações
- Exercício LipídiosDocumento3 páginasExercício LipídiosLayra DandreaAinda não há avaliações
- 03 Peptideos (6 Por Pagina)Documento3 páginas03 Peptideos (6 Por Pagina)Stella Indiukov dos SantosAinda não há avaliações
- Matérias Primas OleaginosasDocumento42 páginasMatérias Primas OleaginosasAdriana Cibele Mesquita DantasAinda não há avaliações
- Hialuroni Relatório de Palavras-Chave Negativas 070222Documento4 páginasHialuroni Relatório de Palavras-Chave Negativas 070222MaumauAinda não há avaliações
- Ficha de Exercicios de Codigo Genetico - Universidade Zambeze - Docx - 1637306221335Documento3 páginasFicha de Exercicios de Codigo Genetico - Universidade Zambeze - Docx - 1637306221335Francisco LourençoAinda não há avaliações
- Biomecânica Básica dos Exercícios: Membros InferioresNo EverandBiomecânica Básica dos Exercícios: Membros InferioresNota: 3.5 de 5 estrelas3.5/5 (8)
- Treinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNo EverandTreinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisNota: 4.5 de 5 estrelas4.5/5 (169)
- Suplementação proteica: Aminoacidemia e saúde vascularNo EverandSuplementação proteica: Aminoacidemia e saúde vascularAinda não há avaliações
- Hackeando Darwin: Engenharia genética e o futuro da humanidadeNo EverandHackeando Darwin: Engenharia genética e o futuro da humanidadeAinda não há avaliações
- Uma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNo EverandUma viagem pelo cérebro: A via rápida para entender neurociência: 1ª edição revisada e atualizadaNota: 4 de 5 estrelas4/5 (13)
- Psicoterapia: Teorias e técnicas psicoterápicasNo EverandPsicoterapia: Teorias e técnicas psicoterápicasNota: 5 de 5 estrelas5/5 (4)
- Aprender mais rápido: Habilidades de estudo, memória e neurociênciaNo EverandAprender mais rápido: Habilidades de estudo, memória e neurociênciaNota: 4 de 5 estrelas4/5 (5)
- Remineralização de Solos com Rochas Ricas em Silício, Potássio e Outros Nutrientes: Uma Solução para Solos Pouco Férteis e Envelhecidos de Cerrados?No EverandRemineralização de Solos com Rochas Ricas em Silício, Potássio e Outros Nutrientes: Uma Solução para Solos Pouco Férteis e Envelhecidos de Cerrados?Ainda não há avaliações
- Tudo tem uma explicação: A biologia por trás de tudo aquilo que você nunca imaginouNo EverandTudo tem uma explicação: A biologia por trás de tudo aquilo que você nunca imaginouNota: 4 de 5 estrelas4/5 (5)