Você também pode gostar
- Treinamento de Corrida de Rua Alexandre Lopes Evangelista - Parte 1Documento10 páginasTreinamento de Corrida de Rua Alexandre Lopes Evangelista - Parte 1matheus ramosAinda não há avaliações
- Fundamentos de BioquímicaDocumento24 páginasFundamentos de BioquímicaCatarina Rodrigues100% (1)
- Nutrição Da SuplementaçãoDocumento28 páginasNutrição Da SuplementaçãoSilvania Santos100% (1)
- Resumo FotossínteseDocumento4 páginasResumo FotossínteseLarissa Feijó100% (1)
- Metabolismo Energético PDFDocumento10 páginasMetabolismo Energético PDFEu Sou BobinhoAinda não há avaliações
- Necessidades Individuais e Necessidades ColetivasDocumento2 páginasNecessidades Individuais e Necessidades ColetivasBraiton antonio100% (1)
- Estudo Sobre A Relação Entre A Fotossíntese e A Respiração Celular Dos VegetaisDocumento14 páginasEstudo Sobre A Relação Entre A Fotossíntese e A Respiração Celular Dos VegetaisCalton AbelAinda não há avaliações
- Aula 9 Degradacao Oxidativa de ProteinasDocumento29 páginasAula 9 Degradacao Oxidativa de ProteinasA Lendária Eguinha PocotóAinda não há avaliações
- Paulo Roberto Silveira - Coletanea de Medicina Ortomolecular III PDFDocumento164 páginasPaulo Roberto Silveira - Coletanea de Medicina Ortomolecular III PDFAnonymous 3a3kkiNgdlAinda não há avaliações
- FOTOSSÍNTESEDocumento20 páginasFOTOSSÍNTESEBruno FelipeAinda não há avaliações
- Ficha Preparação Teste Módulo A3 Com SoluçõesDocumento5 páginasFicha Preparação Teste Módulo A3 Com SoluçõesIsabel Henriques0% (1)
- Ecofisiologia PDFDocumento7 páginasEcofisiologia PDFiristerezinha81Ainda não há avaliações
- Caderno de Aulas Práticas de Patologia para A 1 ProvaDocumento69 páginasCaderno de Aulas Práticas de Patologia para A 1 ProvaVictoria Pagung100% (1)
- Cola P2Documento3 páginasCola P2zepaflaAinda não há avaliações
- Trabalho de Fisiologia NoemiaDocumento14 páginasTrabalho de Fisiologia NoemiaLuís Patrício JoséAinda não há avaliações
- Fisiologia VegetalDocumento3 páginasFisiologia VegetalIsidro Candido Da CostaAinda não há avaliações
- I Experiencia Laboratorial: Verificar A Ocorrência de Fotossíntese e Determinar A Influência Da Luz Na Actividade FotossintéticaDocumento12 páginasI Experiencia Laboratorial: Verificar A Ocorrência de Fotossíntese e Determinar A Influência Da Luz Na Actividade FotossintéticaIssufo SumalgyAinda não há avaliações
- Exercícios de FotossínteseDocumento8 páginasExercícios de FotossínteseJonas MartinsAinda não há avaliações
- Fotossíntese Texto Implicações 2015Documento14 páginasFotossíntese Texto Implicações 2015Nadjama PradoAinda não há avaliações
- Fotoinibi o Da Fotoss NteseDocumento10 páginasFotoinibi o Da Fotoss NteseMa JuhAinda não há avaliações
- Fotossíntese - Resumo, Etapas, Equação, Mapa Mental - Brasil Escola - Reader ViewDocumento8 páginasFotossíntese - Resumo, Etapas, Equação, Mapa Mental - Brasil Escola - Reader ViewMrs. CarrotAinda não há avaliações
- Os Factores Limitantes Do Ecossistema.Documento16 páginasOs Factores Limitantes Do Ecossistema.SergioMatsolo100% (1)
- Rui Baltazar EcossistemaDocumento14 páginasRui Baltazar EcossistemaOdalaAinda não há avaliações
- Aula 1Documento19 páginasAula 1Adriano Silva SouzaAinda não há avaliações
- Metabolismo Do MillhoDocumento16 páginasMetabolismo Do MillhoMomade Ibraimo AssaneAinda não há avaliações
- Seres Autotróficos HiDocumento8 páginasSeres Autotróficos HiInês PereiraAinda não há avaliações
- Fluxo EnergettucoDocumento14 páginasFluxo EnergettucoLoide ManuelAinda não há avaliações
- Curso 23318 Aula 04 Moleculas Celulas e Tecidos v1Documento49 páginasCurso 23318 Aula 04 Moleculas Celulas e Tecidos v1ConcurFernandesAinda não há avaliações
- Fisologia 2Documento3 páginasFisologia 2Queroganhar DinheiroAinda não há avaliações
- W de Manuel - FotossiteseDocumento11 páginasW de Manuel - FotossiteseDeográcio Possiano TalegalAinda não há avaliações
- Trabalho Educacao AmbientalDocumento10 páginasTrabalho Educacao AmbientalFrancisco De Charles CabralAinda não há avaliações
- Trabalho de Fisiologia VegetalDocumento3 páginasTrabalho de Fisiologia VegetalJoaquim Saldeira ManuelAinda não há avaliações
- Fotossintese Aula 4Documento19 páginasFotossintese Aula 4Fernanda BrisonAinda não há avaliações
- Ficha de Leitura - Unidade Tematica IDocumento13 páginasFicha de Leitura - Unidade Tematica IEusebio MuzaluaneAinda não há avaliações
- Ecossistema Do Meu Bairro - QAmbiental - CRCW 2022Documento6 páginasEcossistema Do Meu Bairro - QAmbiental - CRCW 2022Clinton Rafael CaindeAinda não há avaliações
- Fotossíntese ZootecniaDocumento15 páginasFotossíntese ZootecniaMáyra MayAinda não há avaliações
- Semana 10 - Medicina PDFDocumento132 páginasSemana 10 - Medicina PDFKelvyn KennedyAinda não há avaliações
- Quimica Da Atmosfera - Q. Ambiental - CRCW 2022Documento21 páginasQuimica Da Atmosfera - Q. Ambiental - CRCW 2022Clinton Rafael CaindeAinda não há avaliações
- FotossintesebioDocumento13 páginasFotossintesebiolucrencioAinda não há avaliações
- Ciências Da Natureza: FotossínteseDocumento17 páginasCiências Da Natureza: FotossínteseHamfjaj JahfkajdnAinda não há avaliações
- Relação Entre Fotossíntese e Respiração CelularDocumento10 páginasRelação Entre Fotossíntese e Respiração CelularNelson Armando ChilauleAinda não há avaliações
- Reações Luminosas - Aula 5 - Nilo RicardoDocumento7 páginasReações Luminosas - Aula 5 - Nilo RicardoNilo RicardoAinda não há avaliações
- Apontamento 3trimestre - 2021Documento12 páginasApontamento 3trimestre - 2021mchamualiraAinda não há avaliações
- Aula 24 - Química Ambiental: Livro DigitalDocumento158 páginasAula 24 - Química Ambiental: Livro DigitalPaulo RobertoAinda não há avaliações
- FotossínteseDocumento38 páginasFotossínteseArleide NevesAinda não há avaliações
- Botânica FotossínteseDocumento19 páginasBotânica FotossínteseidenildimaAinda não há avaliações
- Biologia-Questionario Fotossintese-Gustavo Moraes Daun-Turma 213 - IFSPDocumento5 páginasBiologia-Questionario Fotossintese-Gustavo Moraes Daun-Turma 213 - IFSPGUSTAVO MORAES DAUNAinda não há avaliações
- Prof Victória Roteiro de Atividades de Ciências - 6º Ano Semana de 26 Á 06 de AgostoDocumento3 páginasProf Victória Roteiro de Atividades de Ciências - 6º Ano Semana de 26 Á 06 de Agostowilian cosmeAinda não há avaliações
- 6 - Fatores Que Afetam FotossínteseeDocumento18 páginas6 - Fatores Que Afetam FotossínteseeHildon Carlos Freitas Rocha FilhoAinda não há avaliações
- Trabalho - 2 de Bioq CatarinaDocumento8 páginasTrabalho - 2 de Bioq Catarinacassimo abilio santosAinda não há avaliações
- Fotossintese IIDocumento12 páginasFotossintese IICleysla LiberalAinda não há avaliações
- Relatório 1Documento19 páginasRelatório 1Carina ArrudaAinda não há avaliações
- Fotossíntese - o Que É, Função, Etapas, Importância - Biologia NetDocumento3 páginasFotossíntese - o Que É, Função, Etapas, Importância - Biologia NetDuda FernandesAinda não há avaliações
- INTERDICIPLINAR (Salvo Automaticamente)Documento12 páginasINTERDICIPLINAR (Salvo Automaticamente)Grasiele AlvesAinda não há avaliações
- Auditoria GruposDocumento80 páginasAuditoria GruposHaznan MichaelaAinda não há avaliações
- Estudo P2 - FotossínteseDocumento9 páginasEstudo P2 - FotossínteseDanili OliveiraAinda não há avaliações
- Trabalho de Biologia - FOTOSSÍNTESEDocumento9 páginasTrabalho de Biologia - FOTOSSÍNTESECínthia GarciaAinda não há avaliações
- Apostil Aula FotossinteseDocumento9 páginasApostil Aula FotossinteseAssessoria AcadêmicaAinda não há avaliações
- Fisiologia VegetalDocumento17 páginasFisiologia VegetalJúpter Horácio AntónioAinda não há avaliações
- Tipos FotossinteseDocumento10 páginasTipos FotossinteseNathália Marcela Batista dos SantosAinda não há avaliações
- Biodiversidade Biológica - ApontamentosDocumento38 páginasBiodiversidade Biológica - Apontamentosmay burgosAinda não há avaliações
- Bioenergetica Top02Documento16 páginasBioenergetica Top02petezbfAinda não há avaliações
- FotossíntesDocumento23 páginasFotossíntesKilson JorgeAinda não há avaliações
- Livro Da Pos 2Documento14 páginasLivro Da Pos 2raulAinda não há avaliações
- Experiências LaboratoriasDocumento13 páginasExperiências Laboratoriasmuchuaneosvaldo4Ainda não há avaliações
- 01 - Influencia Do CO2 Na FotossinteseDocumento14 páginas01 - Influencia Do CO2 Na FotossinteseHeitor CardosoAinda não há avaliações
- Avaliação Projeto Grupo 2Documento7 páginasAvaliação Projeto Grupo 2raul silvaAinda não há avaliações
- Biologia Celular e Molecular: Um guia introdutório para aprendizagem de Biologia Celular e MolecularNo EverandBiologia Celular e Molecular: Um guia introdutório para aprendizagem de Biologia Celular e MolecularAinda não há avaliações
- 30 Autores MoçambicanosDocumento22 páginas30 Autores MoçambicanosBraiton antonioAinda não há avaliações
- Os Princípios Ou Regras Orçamentais 2Documento2 páginasOs Princípios Ou Regras Orçamentais 2Braiton antonioAinda não há avaliações
- Tema de Debate 3Documento3 páginasTema de Debate 3Braiton antonioAinda não há avaliações
- Tema de Debate 2Documento5 páginasTema de Debate 2Braiton antonio100% (1)
- A Interpretação e Integração de Lacunas No Âmbito Da Qualificação e Regime Jurídico Do Direito EmpresarialDocumento1 páginaA Interpretação e Integração de Lacunas No Âmbito Da Qualificação e Regime Jurídico Do Direito EmpresarialBraiton antonioAinda não há avaliações
- Minuta de Pedido de CertificadoDocumento4 páginasMinuta de Pedido de CertificadoBraiton antonio100% (1)
- 1º Trabalho de AtletismoDocumento15 páginas1º Trabalho de AtletismoBraiton antonioAinda não há avaliações
- Aula 07 - Estruturas de Mercado - ConcorrênciaDocumento16 páginasAula 07 - Estruturas de Mercado - ConcorrênciaBraiton antonioAinda não há avaliações
- Estatistica 1Documento4 páginasEstatistica 1Braiton antonioAinda não há avaliações
- Slide Helena Retificado 2023Documento16 páginasSlide Helena Retificado 2023Braiton antonioAinda não há avaliações
- Restaurante JardimDocumento2 páginasRestaurante JardimBraiton antonioAinda não há avaliações
- ProblemaDocumento6 páginasProblemaBraiton antonioAinda não há avaliações
- IntroduçãoDocumento20 páginasIntroduçãoBry PlanetAinda não há avaliações
- A Evolução Da NataçãoDocumento14 páginasA Evolução Da NataçãoIvo CapitulinoAinda não há avaliações
- HelziccccDocumento14 páginasHelziccccBraiton antonioAinda não há avaliações
- Natação Trabalho FeitoDocumento25 páginasNatação Trabalho FeitoBraiton antonioAinda não há avaliações
- Pesquisas em Psicologia e SociologiaDocumento31 páginasPesquisas em Psicologia e SociologiaBraiton antonioAinda não há avaliações
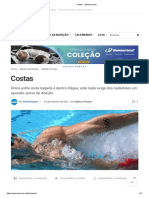
- Costas - SwimchannelDocumento11 páginasCostas - SwimchannelBraiton antonioAinda não há avaliações
- Forum 1Documento4 páginasForum 1Braiton antonioAinda não há avaliações
- Forum 2Documento2 páginasForum 2Braiton antonioAinda não há avaliações
- Relatório - 2022 RetificadoDocumento26 páginasRelatório - 2022 RetificadoBraiton antonioAinda não há avaliações
- Trabalho de FilosofiaDocumento17 páginasTrabalho de FilosofiaBraiton antonioAinda não há avaliações
- Sistema Reprodutivo de Bovino Masculino e FemininoDocumento16 páginasSistema Reprodutivo de Bovino Masculino e FemininoBraiton antonioAinda não há avaliações
- Susana 065329Documento14 páginasSusana 065329Braiton antonioAinda não há avaliações
- Trabalho de Reforma Do Sector PublicoDocumento15 páginasTrabalho de Reforma Do Sector PublicoBraiton antonioAinda não há avaliações
- Trabalho de Contabilidade GeralDocumento17 páginasTrabalho de Contabilidade GeralBraiton antonioAinda não há avaliações
- Trabalho de Antropologia CulturalDocumento14 páginasTrabalho de Antropologia CulturalBraiton antonioAinda não há avaliações
- Trabalho de Psicologia OrganizacionalDocumento13 páginasTrabalho de Psicologia OrganizacionalBraiton antonioAinda não há avaliações
- MIC Trabalho 2 EdsonDocumento20 páginasMIC Trabalho 2 EdsonBraiton antonioAinda não há avaliações
- 1a Fase BiologiaDocumento6 páginas1a Fase BiologiaGabriel SouzaAinda não há avaliações
- Exercícios Bioquímica RevisãoDocumento56 páginasExercícios Bioquímica RevisãoCristafeAinda não há avaliações
- Respiração Celular - TEORIA-IMPORTANTEDocumento5 páginasRespiração Celular - TEORIA-IMPORTANTEDayaneAinda não há avaliações
- Simulado 1/2022 - Ufpr: InstruçõesDocumento35 páginasSimulado 1/2022 - Ufpr: InstruçõesArthur BertoldoAinda não há avaliações
- Biossíntese de Produtos NaturaisDocumento19 páginasBiossíntese de Produtos NaturaisJoao Roberto BeltramoAinda não há avaliações
- LISTA de EXERCÍCIOS - Oxidação de Ácidos Graxos Reg. Met. Ciclo Ureia Corpos CetonicosDocumento6 páginasLISTA de EXERCÍCIOS - Oxidação de Ácidos Graxos Reg. Met. Ciclo Ureia Corpos CetonicosKatia ConceiçãoAinda não há avaliações
- Respiração Aeróbia PDFDocumento2 páginasRespiração Aeróbia PDFGildaMarisaAinda não há avaliações
- Aula 10 - o Ciclo Do Ácido Cítrico p.2Documento24 páginasAula 10 - o Ciclo Do Ácido Cítrico p.2juliana moraes niloAinda não há avaliações
- Glicólise e Ciclo de KrebsDocumento2 páginasGlicólise e Ciclo de Krebspequenalary19Ainda não há avaliações
- Prova Conc. 2016Documento20 páginasProva Conc. 2016Paola Filber100% (1)
- 05 Perito Criminal EspecialDocumento12 páginas05 Perito Criminal EspecialIngride SantosAinda não há avaliações
- Ciclo de Krebs ImprimirDocumento3 páginasCiclo de Krebs ImprimirAna Letícia ReisAinda não há avaliações
- 13 - LipogêneseDocumento4 páginas13 - Lipogênesemaria eduarda100% (1)
- LipídiosDocumento25 páginasLipídiosMaria Rita SalesAinda não há avaliações
- Apostila - Metabolismo EnergéticoDocumento9 páginasApostila - Metabolismo Energéticofiliassa112Ainda não há avaliações
- Pet Eja 1 Ano - 4 BDocumento19 páginasPet Eja 1 Ano - 4 BRosilene Ferreira Araujo100% (1)
- Transformação de Energia Na CélulaDocumento50 páginasTransformação de Energia Na Célulakamilla GarciaAinda não há avaliações
- Ciclo de KrebsDocumento4 páginasCiclo de KrebsDener SilvaAinda não há avaliações
- Apresentação BiologiaDocumento11 páginasApresentação BiologiaAmanda Ketelly Cosme OliveiraAinda não há avaliações
- Respiração Celular e FermentaçãoDocumento7 páginasRespiração Celular e FermentaçãopnrcvvAinda não há avaliações
- Resumo Bioquímica - Glicólise, Ciclo de Krebs, Gliconeogênese, Glicogênese e GlicogenóliseDocumento12 páginasResumo Bioquímica - Glicólise, Ciclo de Krebs, Gliconeogênese, Glicogênese e GlicogenóliseAlexandre DuqueAinda não há avaliações
- 2 Ciclo Do Acido Citrico e Fosforilacao Oxidativa PDFDocumento11 páginas2 Ciclo Do Acido Citrico e Fosforilacao Oxidativa PDFVanessa BispoAinda não há avaliações