Você também pode gostar
- Estudo Dirigido Bioquimica Ciclo de Krebs e Cadeia de Transporte de EletronsDocumento3 páginasEstudo Dirigido Bioquimica Ciclo de Krebs e Cadeia de Transporte de Eletronsadelsonribeiro100% (2)
- 12° ResumoDocumento5 páginas12° ResumoRafaela de Jesus OliveiraAinda não há avaliações
- Tópicos de Estudo - Aula - FODocumento6 páginasTópicos de Estudo - Aula - FOLara SalesAinda não há avaliações
- Cadeia RespiratóriaDocumento60 páginasCadeia RespiratóriaAlline RodriguesAinda não há avaliações
- Visão Geral Da Fosforilação OxidativaDocumento6 páginasVisão Geral Da Fosforilação OxidativaAna Paula AzevedoAinda não há avaliações
- Fosforilação OxidativaDocumento78 páginasFosforilação Oxidativamellogenilza88Ainda não há avaliações
- 20210412-Cadeia Respiratória (Fosforilação Oxidativa) - ADocumento11 páginas20210412-Cadeia Respiratória (Fosforilação Oxidativa) - AISABELA TEIXEIRA DE FREITASAinda não há avaliações
- Processos MitocondriaisDocumento8 páginasProcessos MitocondriaisLone GhostAinda não há avaliações
- Fosforilação OxidativaDocumento36 páginasFosforilação OxidativaDhébora Albuquerque DiasAinda não há avaliações
- Fosforilação OxidativaDocumento36 páginasFosforilação OxidativaCatharina GuimarãesAinda não há avaliações
- Capítulo 19 Do LivroDocumento86 páginasCapítulo 19 Do LivroLeticia LopesAinda não há avaliações
- RESUMO Prova BiocelularDocumento20 páginasRESUMO Prova BiocelularguiarensoAinda não há avaliações
- Respostas BioqDocumento9 páginasRespostas BioqcccaiotestecesarAinda não há avaliações
- Cadeia Transportadora e Fosforilação Oxidativa Retificada-1Documento1 páginaCadeia Transportadora e Fosforilação Oxidativa Retificada-1JohanaAinda não há avaliações
- Bioquímica Metabólica 02Documento26 páginasBioquímica Metabólica 02Viviane RangelAinda não há avaliações
- 369785-Fosforilação OxidativaDocumento4 páginas369785-Fosforilação OxidativaRamires Cerqueira100% (1)
- Fosforolização OxidativaDocumento16 páginasFosforolização Oxidativabrunowarley1982Ainda não há avaliações
- AULA 1 - Respiração CelularDocumento75 páginasAULA 1 - Respiração CelularWilliams Fabi KawanyAinda não há avaliações
- Fosforilação OxidativaDocumento31 páginasFosforilação Oxidativatitalves100% (1)
- Aula Metabolismo Celular - RespiraçãoDocumento29 páginasAula Metabolismo Celular - RespiraçãoLúcio Eduardo FerreiraAinda não há avaliações
- Fosforilação Oxidativa e Cadeia Transportadora de ElétronsDocumento42 páginasFosforilação Oxidativa e Cadeia Transportadora de ElétronsFernando BritoAinda não há avaliações
- Estudo P2 - BioquímicaDocumento20 páginasEstudo P2 - BioquímicaMirella FrancoAinda não há avaliações
- Cadeia Transportadora de ElétronsDocumento3 páginasCadeia Transportadora de ElétronsMaria Eduarda DiasAinda não há avaliações
- Aula 041599763689Documento15 páginasAula 041599763689SGI Gestão IntegradoAinda não há avaliações
- 369784-Cadeia de Transporte de ElétronsDocumento4 páginas369784-Cadeia de Transporte de ElétronsRamires CerqueiraAinda não há avaliações
- Estudo Dirigido - Mitocôndrias e Cloroplastos.Documento3 páginasEstudo Dirigido - Mitocôndrias e Cloroplastos.Luiza LopesAinda não há avaliações
- Anotações BioquímicaDocumento10 páginasAnotações BioquímicaAnderson Souza de AssisAinda não há avaliações
- Aula 7 - Fosforilacao OxidativaDocumento32 páginasAula 7 - Fosforilacao OxidativaEveny PereiraAinda não há avaliações
- Aula - RESPIRAÇÃO CELULAR - Parte III - Fosforilação OxidativaDocumento23 páginasAula - RESPIRAÇÃO CELULAR - Parte III - Fosforilação OxidativaAntonio Carlos Silva-JúniorAinda não há avaliações
- Slides Cadeia Transportadora de EletronsDocumento12 páginasSlides Cadeia Transportadora de EletronscelsogdjAinda não há avaliações
- Formação de Atp Nas MitocondriasDocumento2 páginasFormação de Atp Nas MitocondriasThalitaAinda não há avaliações
- GD Transporte ResolvidoDocumento6 páginasGD Transporte ResolvidoSheila PinhoAinda não há avaliações
- Estudo Dirigido - ClotoplastosDocumento3 páginasEstudo Dirigido - ClotoplastosLuiza LopesAinda não há avaliações
- Bioenergética-Mitocôndrias e Respiração CelularDocumento11 páginasBioenergética-Mitocôndrias e Respiração CelularErica Rosa RangelAinda não há avaliações
- Exercicios Terceiro ModuloDocumento3 páginasExercicios Terceiro ModuloGabriela AndradeAinda não há avaliações
- BIO10 - PPT - D4 - 1. Obtenção de EnergiaDocumento22 páginasBIO10 - PPT - D4 - 1. Obtenção de Energiacaroltdsousa02Ainda não há avaliações
- Bioinorgnica 1 - Fontes de Energia e FunesDocumento49 páginasBioinorgnica 1 - Fontes de Energia e FunesNathália CarvalhoAinda não há avaliações
- Fosforilação Oxidativa SlidesDocumento23 páginasFosforilação Oxidativa SlidesAniliowedy greatAinda não há avaliações
- Correcção Das Perguntas de BiologiaDocumento4 páginasCorrecção Das Perguntas de BiologiaTomás D'Azevedo LopesAinda não há avaliações
- Resumo - Fosforilacao OxidativaDocumento3 páginasResumo - Fosforilacao OxidativaIvan AlvesAinda não há avaliações
- Respiração CelularDocumento22 páginasRespiração CelularBruno FelipeAinda não há avaliações
- Slides Aula 11. CTE e Fosforilação OxidativaDocumento26 páginasSlides Aula 11. CTE e Fosforilação OxidativaÂngelo AbnerAinda não há avaliações
- Ciclo de Krebs e Cadeia Transportadora de ElétronsDocumento2 páginasCiclo de Krebs e Cadeia Transportadora de ElétronsLuna NavarroAinda não há avaliações
- Metabolismo MicrobianoDocumento7 páginasMetabolismo Microbianograsiela camposAinda não há avaliações
- Estudo Dirigido 3 - Bioenergética e CTE - Com RESPOSTASDocumento4 páginasEstudo Dirigido 3 - Bioenergética e CTE - Com RESPOSTASblimaloveAinda não há avaliações
- Resumo BioquA - Mica II - P2Documento21 páginasResumo BioquA - Mica II - P2Caroline Carvalho LimaAinda não há avaliações
- Fosforilacao OxidativaDocumento24 páginasFosforilacao OxidativaLucas Roniery100% (1)
- (Alberts, 4 Ed) Cap. 14 - Geração de Energia Nas MitocôndriasDocumento8 páginas(Alberts, 4 Ed) Cap. 14 - Geração de Energia Nas MitocôndriasDenis NascimentoAinda não há avaliações
- Estudo DirigidoDocumento5 páginasEstudo DirigidoLaianaCarmanini100% (2)
- CIENCIA HOJE Vol 34 N 199 MitocondriaDocumento8 páginasCIENCIA HOJE Vol 34 N 199 MitocondriaLeticia EmidioAinda não há avaliações
- A Cadeia de Transporte de ElétronsDocumento8 páginasA Cadeia de Transporte de ElétronsSamilly BarbosaAinda não há avaliações
- Metabolismo de CH (Apostila)Documento5 páginasMetabolismo de CH (Apostila)Fernanda LacerdaAinda não há avaliações
- Cadeia Transportadora de Elétrons e Fosforilação Oxidativa PDFDocumento48 páginasCadeia Transportadora de Elétrons e Fosforilação Oxidativa PDFDiesSyca Masson MassonzinhaAinda não há avaliações
- Prova 2 BioquímicaDocumento10 páginasProva 2 BioquímicaKyara De JulioAinda não há avaliações
- Fosforilação Oxidativa CompletaDocumento27 páginasFosforilação Oxidativa CompletaLuiz Humberto Souza JúniorAinda não há avaliações
- Wa0001Documento12 páginasWa0001kokmerpqAinda não há avaliações
- 004 Aula Semana 4Documento37 páginas004 Aula Semana 4Gabriella S. de SouzaAinda não há avaliações
- Respiração - Celular Aula 06Documento27 páginasRespiração - Celular Aula 06Fernando Alves CostaAinda não há avaliações
- 13,14 e 15 OxidaçãoDocumento1 página13,14 e 15 OxidaçãoGuilherme Belak SchmitkeAinda não há avaliações
- Lista Exercícios - Carboidratos e LipídeosDocumento10 páginasLista Exercícios - Carboidratos e LipídeosGuilherme Belak SchmitkeAinda não há avaliações
- Atividade Eletro 5Documento2 páginasAtividade Eletro 5Guilherme Belak SchmitkeAinda não há avaliações
- FIBRASDocumento10 páginasFIBRASGuilherme Belak SchmitkeAinda não há avaliações
- Lista de Exercícios - EnzimasDocumento5 páginasLista de Exercícios - EnzimasGuilherme Belak SchmitkeAinda não há avaliações
- 11 e 12 OxidaçãoDocumento1 página11 e 12 OxidaçãoGuilherme Belak SchmitkeAinda não há avaliações
- FÓRMULAS DRIs PARA CÁLCULO DO GEB E DO GET 24.05.17Documento8 páginasFÓRMULAS DRIs PARA CÁLCULO DO GEB E DO GET 24.05.17ROSANGELA APARECIDA XIMENEZAinda não há avaliações
- Atividade Eletro 3Documento25 páginasAtividade Eletro 3Guilherme Belak SchmitkeAinda não há avaliações
- Tabela Per CapitasDocumento5 páginasTabela Per CapitasGuilherme Belak SchmitkeAinda não há avaliações
- Manejo Do LixoDocumento1 páginaManejo Do LixoGuilherme Belak SchmitkeAinda não há avaliações
- Quando Os Micro Se MultiplicamDocumento1 páginaQuando Os Micro Se MultiplicamGuilherme Belak SchmitkeAinda não há avaliações
- Plano de Aula - RadioatividadeDocumento3 páginasPlano de Aula - RadioatividadeGuilherme Belak SchmitkeAinda não há avaliações
- Fluxograma Descongelamento de AlimentosDocumento1 páginaFluxograma Descongelamento de AlimentosGuilherme Belak SchmitkeAinda não há avaliações
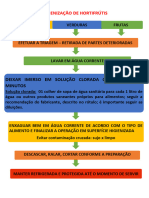
- Fluxograma Higienização de HortifrutisDocumento2 páginasFluxograma Higienização de HortifrutisGuilherme Belak SchmitkeAinda não há avaliações
- Plano de Aula Modelos AtomicosDocumento2 páginasPlano de Aula Modelos AtomicosGuilherme Belak SchmitkeAinda não há avaliações
- Plano de Aula - Dia Consc. Negra - Funções OrgânicasDocumento3 páginasPlano de Aula - Dia Consc. Negra - Funções OrgânicasGuilherme Belak SchmitkeAinda não há avaliações
- Plano de Aula Modelos AtomicosDocumento2 páginasPlano de Aula Modelos AtomicosGuilherme Belak SchmitkeAinda não há avaliações
- AlotropiaPlano de AulaDocumento2 páginasAlotropiaPlano de AulaGuilherme Belak SchmitkeAinda não há avaliações
- Plano de Aula Modelos AtomicosDocumento2 páginasPlano de Aula Modelos AtomicosGuilherme Belak SchmitkeAinda não há avaliações
- Plano de Aula - Dia Consc. Negra - Misturas e Processos de SeparaçãoDocumento3 páginasPlano de Aula - Dia Consc. Negra - Misturas e Processos de SeparaçãoGuilherme Belak SchmitkeAinda não há avaliações
- Teoria Do Canal Aplicado Na MTC - PortuguêsDocumento756 páginasTeoria Do Canal Aplicado Na MTC - PortuguêsWEROTTYDESAinda não há avaliações
- 10 Passos para Um Evangelismo Pessoal EficienteDocumento2 páginas10 Passos para Um Evangelismo Pessoal EficienteWelhington S. da SilvaAinda não há avaliações
- Recepção Dos Conceitos AgostinianosDocumento193 páginasRecepção Dos Conceitos AgostinianosWallace Johnson100% (1)
- Unidade 1 - AmoniacoDocumento40 páginasUnidade 1 - Amoniacoantónio_ramalho_17Ainda não há avaliações
- Oracao de Sao BentoDocumento1 páginaOracao de Sao BentoEduardo Luis SouzaAinda não há avaliações
- Direito Penal IDocumento57 páginasDireito Penal Icarina nunesAinda não há avaliações
- INTERNACIONAL SEMANA 11 - RespostaDocumento2 páginasINTERNACIONAL SEMANA 11 - RespostaAdenilson Nascimento SoaresAinda não há avaliações
- O Asno, A Raposa e o LeãoDocumento3 páginasO Asno, A Raposa e o LeãoSonicGeneraçãoAinda não há avaliações
- Os HOPI também acreditam na emergência e extinção cíclica dos Homens, que se renovam em raças cada vez mais evoluídas rumo a uma purificação espiritual que chegará ao termo ideal na Sétima Raça ou Sétimo MundDocumento5 páginasOs HOPI também acreditam na emergência e extinção cíclica dos Homens, que se renovam em raças cada vez mais evoluídas rumo a uma purificação espiritual que chegará ao termo ideal na Sétima Raça ou Sétimo MundMarcos CabralAinda não há avaliações
- Atividade Portugueses Na AméricaDocumento1 páginaAtividade Portugueses Na AméricaKatrine Antunes PachecoAinda não há avaliações
- Prova 2018 para C-FSD-FN 2019Documento7 páginasProva 2018 para C-FSD-FN 2019pepu mpAinda não há avaliações
- Aula 5 - Mapeamento Do Modelo Entidade Relacionamento para o Modelo RelacionalDocumento2 páginasAula 5 - Mapeamento Do Modelo Entidade Relacionamento para o Modelo RelacionalJoao SilvaAinda não há avaliações
- Síntese A Reforma Sanitária e o Processo de Implantação Do Sistema Único de Saúde No BrasilDocumento3 páginasSíntese A Reforma Sanitária e o Processo de Implantação Do Sistema Único de Saúde No BrasilGabriel PortoAinda não há avaliações
- Relatório Extração Solido LiquidoDocumento15 páginasRelatório Extração Solido LiquidoJessyka NicollyAinda não há avaliações
- Psicologia e ReligiãoDocumento7 páginasPsicologia e ReligiãoFranciscoWalissonAinda não há avaliações
- Sintonia Perfeita (Serie The Re - Amanda Maia-1Documento951 páginasSintonia Perfeita (Serie The Re - Amanda Maia-1Sofia Almeida100% (2)
- Dermatoses PsicogenicasDocumento80 páginasDermatoses PsicogenicasCláudia Da Costa SilvaAinda não há avaliações
- Apostila Do Modulo 3 - Empreendedorismo PDFDocumento27 páginasApostila Do Modulo 3 - Empreendedorismo PDFArthemia SampaioAinda não há avaliações
- Aula 7 - ToxicodinâmicaDocumento39 páginasAula 7 - ToxicodinâmicaFernanda MoreiraAinda não há avaliações
- Religiões e Modernidades: Cristianismos, Secularização e Novas EspiritualidadesDocumento271 páginasReligiões e Modernidades: Cristianismos, Secularização e Novas EspiritualidadesEditora Pimenta CulturalAinda não há avaliações
- 01 o Pecado OriginalDocumento42 páginas01 o Pecado OriginalNelson FerreiraAinda não há avaliações
- Febrapils - Código de Conduta e Ética (2014-17)Documento5 páginasFebrapils - Código de Conduta e Ética (2014-17)José Flávio da Paz100% (1)
- Relatório Ensaio de DurezaDocumento13 páginasRelatório Ensaio de DurezaIsabelle BarachoAinda não há avaliações
- Atividade Objetiva 4 - Antropologia - Identidade e DiversidadeDocumento10 páginasAtividade Objetiva 4 - Antropologia - Identidade e DiversidadeFabiola RodriguesAinda não há avaliações
- BG11 Teste Evol Classif 2011Documento5 páginasBG11 Teste Evol Classif 2011Ana Fonte100% (1)
- 019 AULA 07 Didática Planejamento e Avaliação 1Documento18 páginas019 AULA 07 Didática Planejamento e Avaliação 1João Brunno SucupiraAinda não há avaliações
- Iconicidade e ArbitrariedadeDocumento2 páginasIconicidade e ArbitrariedadeMauricio Damasceno SouzaAinda não há avaliações
- Proposiciones y Sus Contracciones - Tus Clases de PortuguésDocumento4 páginasProposiciones y Sus Contracciones - Tus Clases de PortuguésCarmen Talledo GalvezAinda não há avaliações
- Catalogo GrafismosDocumento33 páginasCatalogo Grafismosanaemidiasousarocha100% (2)
- Nutrimentos Parte 3 PDFDocumento18 páginasNutrimentos Parte 3 PDFlauryAinda não há avaliações