Você também pode gostar
- Sistema imunológico: células, órgãos e respostas inatas e adaptativasDocumento109 páginasSistema imunológico: células, órgãos e respostas inatas e adaptativasIngridAinda não há avaliações
- Imuno IIDocumento26 páginasImuno IIJonatan Dos Santos De OliveiraAinda não há avaliações
- Imunidade Inata: Primeira Linha de DefesaDocumento63 páginasImunidade Inata: Primeira Linha de DefesaNina Maria100% (1)
- Imunologia Basica 6ed 2021 AbbasDocumento69 páginasImunologia Basica 6ed 2021 AbbasCarina RibeiroAinda não há avaliações
- Avanços da biologia celular e da genética molecularNo EverandAvanços da biologia celular e da genética molecularAinda não há avaliações
- Do comensalismo à patogenicidade: Um estudo sobre estafilococos coagulase-negativaNo EverandDo comensalismo à patogenicidade: Um estudo sobre estafilococos coagulase-negativaAinda não há avaliações
- Enterobactérias: taxonomia, características e doenças causadasDocumento13 páginasEnterobactérias: taxonomia, características e doenças causadaslucianofloresAinda não há avaliações
- Taxonomia MicrobiológicaDocumento23 páginasTaxonomia MicrobiológicaVanessa LimaAinda não há avaliações
- Sistema complemento principal defesa inataDocumento23 páginasSistema complemento principal defesa inataThayseVazAinda não há avaliações
- Morte celular: necrose e apoptoseDocumento34 páginasMorte celular: necrose e apoptoseEdson LuccasAinda não há avaliações
- Diversidade Dos Anticorpos Imunologia BasicaDocumento46 páginasDiversidade Dos Anticorpos Imunologia BasicaJonas SaldanhaAinda não há avaliações
- Tricuríase e EnterobioseDocumento5 páginasTricuríase e EnterobioseMilhoeAinda não há avaliações
- Aula 6 Sistema ComplementoDocumento27 páginasAula 6 Sistema ComplementoDanielly BogeaAinda não há avaliações
- Microbiologia Aula VirusDocumento6 páginasMicrobiologia Aula Viruswillem douglas100% (1)
- Medicina Veterinária Investigativa (Patologia Veterinária) - ResumoDocumento9 páginasMedicina Veterinária Investigativa (Patologia Veterinária) - ResumoKauã SantosAinda não há avaliações
- Micologia: Estudo dos fungosDocumento48 páginasMicologia: Estudo dos fungosWalter MoreiraAinda não há avaliações
- E. coli Características Patogênese DoençasDocumento15 páginasE. coli Características Patogênese DoençasPaula CarvalhoAinda não há avaliações
- Aula 04 - ImunologiaDocumento20 páginasAula 04 - ImunologiaJairlane Garcia100% (1)
- Imunologia - Anticorpos e VacinasDocumento13 páginasImunologia - Anticorpos e VacinasJanyne Feitosa100% (1)
- Sistema complemento - Funções e vias de ativaçãoDocumento11 páginasSistema complemento - Funções e vias de ativaçãoGerson Coelho VianaAinda não há avaliações
- Lista de Exercícios de Biologia MolecularDocumento2 páginasLista de Exercícios de Biologia MolecularAnderson MatheusAinda não há avaliações
- Aula HPA3 CarrapatosDocumento69 páginasAula HPA3 CarrapatosJoao Vitor Pereira Marques BorgesAinda não há avaliações
- Introdução À BacteriologiaDocumento91 páginasIntrodução À BacteriologiaMelissa ArraesAinda não há avaliações
- 1 HematologiaDocumento41 páginas1 HematologiaAna Cláudia SouzaAinda não há avaliações
- Resumo final Microbiologia I: principais conceitos e métodosDocumento31 páginasResumo final Microbiologia I: principais conceitos e métodosMichelle CristineAinda não há avaliações
- Diagnósticos laboratoriais em Parasitologia ClínicaDocumento28 páginasDiagnósticos laboratoriais em Parasitologia Clínicaduilio junior100% (1)
- Estudo sobre Parasitologia com 15 questõesDocumento3 páginasEstudo sobre Parasitologia com 15 questõesIZABELA DA ROCHA ARRUDAAinda não há avaliações
- Sistema ComplementoDocumento19 páginasSistema ComplementoJanaína Andrade100% (1)
- PlatyhelminthesDocumento18 páginasPlatyhelminthesangel ber0% (1)
- Transtornos ImunológicosDocumento53 páginasTranstornos ImunológicosEdnilson Jordao MassicameAinda não há avaliações
- Antígeno e anticorpo: estrutura, funções e diversidadeDocumento53 páginasAntígeno e anticorpo: estrutura, funções e diversidadeRodrygo Arruda100% (1)
- Imunologia Dos Tumores SlideDocumento23 páginasImunologia Dos Tumores SlideBiomedicina CeumaAinda não há avaliações
- Micotoxinas na produção suínaDocumento89 páginasMicotoxinas na produção suínaSicelino MeiraAinda não há avaliações
- Malria PDFDocumento23 páginasMalria PDFAlex BritoAinda não há avaliações
- Aula 16 OncocercoseDocumento20 páginasAula 16 Oncocercosemanoel alencarAinda não há avaliações
- Apostila de Imunologia (Pratica)Documento100 páginasApostila de Imunologia (Pratica)yurico122Ainda não há avaliações
- Introdução à Imunologia VeterináriaDocumento10 páginasIntrodução à Imunologia Veterinárialarissa doppus0% (1)
- Aula 06 - BabesiidaeDocumento41 páginasAula 06 - BabesiidaeGabrieli MolinaAinda não há avaliações
- Coleta, Armazenamento e Envio de Amostras de Material de Aborto para Diagnóstico de Doenças Infecciosas em BovinosDocumento6 páginasColeta, Armazenamento e Envio de Amostras de Material de Aborto para Diagnóstico de Doenças Infecciosas em BovinosjaimecafigueroaAinda não há avaliações
- Mielograma: indicações e interpretaçãoDocumento95 páginasMielograma: indicações e interpretaçãodouglas648Ainda não há avaliações
- Imunoensaio RápidoDocumento26 páginasImunoensaio RápidoMarianaAinda não há avaliações
- Teníase e Cisticercose: transmissão, sinais e prevençãoDocumento20 páginasTeníase e Cisticercose: transmissão, sinais e prevençãoJeremy S. FölderAinda não há avaliações
- Roteiro de Estudo para Prova Prática de MicologiaDocumento5 páginasRoteiro de Estudo para Prova Prática de Micologiakamila123nunesAinda não há avaliações
- Replicacao Viral 2012 BiomedDocumento32 páginasReplicacao Viral 2012 BiomedFabiane RibeiroAinda não há avaliações
- Resumo ImunohematoDocumento9 páginasResumo ImunohematolittlebruAinda não há avaliações
- Sistema circulatório e componentes do sangueDocumento50 páginasSistema circulatório e componentes do sangueCarlos Renato NogueiraAinda não há avaliações
- (IMUNO) Imunologia Antígeno-Anticorpo - RESUMODocumento9 páginas(IMUNO) Imunologia Antígeno-Anticorpo - RESUMOSamara SantosAinda não há avaliações
- Ciclos de Trypanosoma, Leishmania, Toxoplasma e PlasmodiumDocumento7 páginasCiclos de Trypanosoma, Leishmania, Toxoplasma e PlasmodiumNathália SilvaAinda não há avaliações
- Imunidade e RespostasDocumento9 páginasImunidade e RespostasRenato RibeiroAinda não há avaliações
- Hemoparasitose - Aula 7Documento12 páginasHemoparasitose - Aula 7jessicaAinda não há avaliações
- Apostila de Bacteriologia Médica 2015-1Documento111 páginasApostila de Bacteriologia Médica 2015-1Ana Clara Rocha0% (1)
- A vida é um ciclo onde o princípio e o fim se unemDocumento18 páginasA vida é um ciclo onde o princípio e o fim se unemDalila CaixetaAinda não há avaliações
- ELISA: Enzyme-Linked Immunosorbent AssayDocumento10 páginasELISA: Enzyme-Linked Immunosorbent AssayLuisa SixxAinda não há avaliações
- Conceitos em ImunologiaDocumento39 páginasConceitos em ImunologiaRafaela BritoAinda não há avaliações
- Pitiose EquinaDocumento9 páginasPitiose EquinaTony PicoliAinda não há avaliações
- Filo Nematoda: morfologia, ciclo de vida e parasitas importantesDocumento37 páginasFilo Nematoda: morfologia, ciclo de vida e parasitas importantesSandy LorrayneAinda não há avaliações
- Aula Introdução Ao Imunodiagnóstico OKDocumento45 páginasAula Introdução Ao Imunodiagnóstico OKOdilon Calian100% (1)
- BabesioseDocumento29 páginasBabesioseVictoria RomanoAinda não há avaliações
- 6 Ativação de Linfocito TDocumento11 páginas6 Ativação de Linfocito TLeticia CostaAinda não há avaliações
- Ativação de linfócitos B: maturação, fases da resposta e reconhecimento de antígenosDocumento11 páginasAtivação de linfócitos B: maturação, fases da resposta e reconhecimento de antígenosLeticia CostaAinda não há avaliações
- 1 Tecidos LinfoidesDocumento8 páginas1 Tecidos LinfoidesLeticia CostaAinda não há avaliações
- 4 AnticorpoDocumento6 páginas4 AnticorpoLeticia CostaAinda não há avaliações
- Cardiopatias Congênitas AcianóticasDocumento15 páginasCardiopatias Congênitas AcianóticasLeticia CostaAinda não há avaliações
- Definição, classificação e conduta na diarreia aguda e crônicaDocumento11 páginasDefinição, classificação e conduta na diarreia aguda e crônicaLeticia CostaAinda não há avaliações
- Anemias - Ferropriva, B12, Aplásica e FalciformeDocumento15 páginasAnemias - Ferropriva, B12, Aplásica e FalciformeLeticia CostaAinda não há avaliações
- 21019f-Diretrizes Consenso Sobre Anemia Ferropriva-OkDocumento13 páginas21019f-Diretrizes Consenso Sobre Anemia Ferropriva-OkCarloJamaAinda não há avaliações
- Anemia ferropriva: atualização das diretrizes de 2021Documento7 páginasAnemia ferropriva: atualização das diretrizes de 2021Laura SabrinaAinda não há avaliações
- Sodio - Crise Hipert - Page 1Documento1 páginaSodio - Crise Hipert - Page 1Leticia CostaAinda não há avaliações
- MeningiteDocumento4 páginasMeningiteLeticia CostaAinda não há avaliações
- Choque PediatriaDocumento1 páginaChoque PediatriaLeticia CostaAinda não há avaliações
- Dçs Tubulo IntersticiaisDocumento2 páginasDçs Tubulo IntersticiaisLeticia CostaAinda não há avaliações
- SÍNDROME CORONARIANA AGUDADocumento1 páginaSÍNDROME CORONARIANA AGUDALeticia CostaAinda não há avaliações
- Hipercalemia: alterações ECG e tratamentoDocumento2 páginasHipercalemia: alterações ECG e tratamentoLeticia CostaAinda não há avaliações
- Causas e Características da Glomerulonefrite AgudaDocumento7 páginasCausas e Características da Glomerulonefrite AgudaLeticia CostaAinda não há avaliações
- Membrana Celular - Composição e FunçõesDocumento9 páginasMembrana Celular - Composição e FunçõesElder Andrade AmaralAinda não há avaliações
- Imunologia Resumida - 2 Ed.Documento140 páginasImunologia Resumida - 2 Ed.Luís GustavoAinda não há avaliações
- Captura de Tela 2024-02-19 À(s) 09.01.26Documento21 páginasCaptura de Tela 2024-02-19 À(s) 09.01.2665z8spfjtdAinda não há avaliações
- BioeletrogêneseDocumento7 páginasBioeletrogêneseCarol RégisAinda não há avaliações
- Lista de Exercícios ImunologiaDocumento4 páginasLista de Exercícios ImunologiaIago GabrielAinda não há avaliações
- Células Do Sistema ImuneDocumento6 páginasCélulas Do Sistema ImuneLaysa Gomes dos SantosAinda não há avaliações
- Plano de ensino de Imunologia Básica para BiomedicinaDocumento3 páginasPlano de ensino de Imunologia Básica para BiomedicinaAline Preve da SilvaAinda não há avaliações
- Fichamento Dinâmica Da Resposta Imune ImunologiaDocumento1 páginaFichamento Dinâmica Da Resposta Imune ImunologiaJulia Lopes MouraAinda não há avaliações
- Celulas Sistema ImuneDocumento135 páginasCelulas Sistema ImuneMiriam NutricionistaAinda não há avaliações
- Escopolamina Golan Farmacologia Capitulo 08Documento19 páginasEscopolamina Golan Farmacologia Capitulo 08Leandro Freneda100% (1)
- Aula 2 - Funcionamento Geral Do Sistema ImunológicoDocumento34 páginasAula 2 - Funcionamento Geral Do Sistema ImunológicoDavid M SampaioAinda não há avaliações
- Potenciais de MembranaDocumento36 páginasPotenciais de MembranaLuidi Marques100% (1)
- Sistema de endomembranasDocumento3 páginasSistema de endomembranasDaniela BarbosaAinda não há avaliações
- Estudo Dirigido - Bioquimica ClinicaDocumento2 páginasEstudo Dirigido - Bioquimica ClinicaIsabela Coimbra Ramos100% (1)
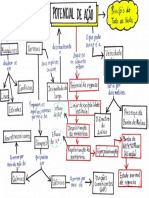
- Mapa Conceitual - Potencial de AçãoDocumento1 páginaMapa Conceitual - Potencial de AçãoLarissa LimaAinda não há avaliações
- Sistema ComplementoDocumento40 páginasSistema ComplementoMillena CoelhoAinda não há avaliações
- Resumo CONFICT 2023 - Ana Luiza - FINALDocumento2 páginasResumo CONFICT 2023 - Ana Luiza - FINALAna Luiza Moreira do Nascimento ValenteAinda não há avaliações
- Biofísica das membranas excitáveis e BioeletrogêneseDocumento30 páginasBiofísica das membranas excitáveis e BioeletrogêneseBarbara GranatoAinda não há avaliações
- Sistemas Fisiologia HumanaDocumento2 páginasSistemas Fisiologia Humanaprof.analuciaAinda não há avaliações
- Introdução à ImunologiaDocumento4 páginasIntrodução à ImunologiaLeonardo Oliveira100% (1)
- Receptores Acoplados À Proteína GDocumento3 páginasReceptores Acoplados À Proteína Graianna_17Ainda não há avaliações
- Aula 04 - Sistema ComplementoDocumento19 páginasAula 04 - Sistema Complementopamelacunhaenf1Ainda não há avaliações
- Atividade de Imunologia - Docx Atividade 2Documento3 páginasAtividade de Imunologia - Docx Atividade 2Joyce SilvaAinda não há avaliações
- AR - Celulas B - 230427 - 105931Documento14 páginasAR - Celulas B - 230427 - 105931Larissa HayneAinda não há avaliações
- Digestão e Absorção de ProteínasDocumento20 páginasDigestão e Absorção de ProteínasBoris Curioso CuriosoAinda não há avaliações
- CRISTALOGRAFIA - UnBDocumento3 páginasCRISTALOGRAFIA - UnBRamon AlbuquerqueAinda não há avaliações
- Imunidade inata: componentes e funçõesDocumento33 páginasImunidade inata: componentes e funçõesVictoria MendesAinda não há avaliações
- Membrana Plasmática 1Documento27 páginasMembrana Plasmática 1Ines CostaAinda não há avaliações
- Processo de maturação dos linfócitos B e TDocumento3 páginasProcesso de maturação dos linfócitos B e TJúlio CésarAinda não há avaliações
- Membrana PlasmáticaDocumento22 páginasMembrana Plasmáticaericmhara23Ainda não há avaliações