Você também pode gostar
- Estudo Dirigido BioquimicaDocumento5 páginasEstudo Dirigido Bioquimicaadelsonribeiro100% (4)
- Resumo - GlicóliseDocumento11 páginasResumo - Glicólisemaelson100% (2)
- Questões Respondidas Sobre A 2 e 3 Prova de BioquímicaDocumento40 páginasQuestões Respondidas Sobre A 2 e 3 Prova de BioquímicaViviane Moura60% (5)
- LIsta Met GlicogênioDocumento4 páginasLIsta Met GlicogênioAlbert DiasAinda não há avaliações
- Topologia dos Principais Transportadores de Açúcares: uma abordagem sobre a estrutura e função dos GLUT'sNo EverandTopologia dos Principais Transportadores de Açúcares: uma abordagem sobre a estrutura e função dos GLUT'sAinda não há avaliações
- Tabela Per CapitasDocumento5 páginasTabela Per CapitasGuilherme Belak SchmitkeAinda não há avaliações
- NBR 15515-3 - DetalhadaDocumento22 páginasNBR 15515-3 - DetalhadaHoracimar CotrimAinda não há avaliações
- Metabolismo Dos HC - Glicogénsese e Glicogenólise 05-11-2018Documento28 páginasMetabolismo Dos HC - Glicogénsese e Glicogenólise 05-11-2018DiogoFevereiroAinda não há avaliações
- Cap 11 - Metabolismo Do GlicogênioDocumento3 páginasCap 11 - Metabolismo Do Glicogêniojbod2012100% (1)
- Biossintese de Carboidratos BioquímicaDocumento6 páginasBiossintese de Carboidratos BioquímicaMarcela Lima de BritoAinda não há avaliações
- Resposta Estudo Dirigido BqiiDocumento13 páginasResposta Estudo Dirigido BqiiVitor Ferreira ValenteAinda não há avaliações
- Resumos BioquímicaDocumento14 páginasResumos BioquímicaGabrieli HoepersAinda não há avaliações
- Metabolismo Do GlicognioDocumento15 páginasMetabolismo Do GlicognioDaniel FreireAinda não há avaliações
- Aula 10 - Glicogenese GlicogenoliseDocumento37 páginasAula 10 - Glicogenese GlicogenoliseGabriela SteimbachAinda não há avaliações
- Metabolismo de CH II (Odonto)Documento17 páginasMetabolismo de CH II (Odonto)Fernanda LacerdaAinda não há avaliações
- Metabolismo EnergéticoDocumento7 páginasMetabolismo EnergéticoMariana PaesAinda não há avaliações
- Metabolismo de CarboidratosDocumento4 páginasMetabolismo de CarboidratosHellycláudia ChavesAinda não há avaliações
- Aula 14 - GluconeogéneseDocumento24 páginasAula 14 - GluconeogéneseaptgaptgAinda não há avaliações
- 6.2 Metabolismo de CHO - Gliconeogênese e GlicogênioDocumento20 páginas6.2 Metabolismo de CHO - Gliconeogênese e GlicogênioAna SilvaAinda não há avaliações
- Capítulo 14 Lehninger + ExtrasDocumento60 páginasCapítulo 14 Lehninger + ExtrasDaniel FreireAinda não há avaliações
- BioquimiscDocumento7 páginasBioquimiscJuca Do sabugueiro loucoAinda não há avaliações
- Metabolismo Do GlicogênioDocumento3 páginasMetabolismo Do GlicogênioLuanaAinda não há avaliações
- Glicolise 05082010Documento57 páginasGlicolise 05082010andre_iaiaAinda não há avaliações
- Lista GliconeogeneseDocumento6 páginasLista GliconeogeneseGuilherme Belak SchmitkeAinda não há avaliações
- 11284816022012bioquimica Aula 13 PDFDocumento18 páginas11284816022012bioquimica Aula 13 PDFDiego PereiraAinda não há avaliações
- Metabolismo Do GlicogênioDocumento6 páginasMetabolismo Do GlicogênioSara EsterAinda não há avaliações
- 6 - Metabolismo de Carboidratos I - Glicólise e GliconeogêneseDocumento8 páginas6 - Metabolismo de Carboidratos I - Glicólise e GliconeogêneseFelipe RostirollaAinda não há avaliações
- Atividade GliconeogêneseDocumento5 páginasAtividade GliconeogêneseGatodo FabaAinda não há avaliações
- Lista de Exercicios Aula 13-Ciclo de Krebs e GlicogênioDocumento7 páginasLista de Exercicios Aula 13-Ciclo de Krebs e GlicogênioMaria MoreiraAinda não há avaliações
- Aula03 BioqII-CFBio Glioconeogenese1Documento38 páginasAula03 BioqII-CFBio Glioconeogenese1hide-seekAinda não há avaliações
- Uc3 P2Documento10 páginasUc3 P2Lucas KallielAinda não há avaliações
- Gliconeogenese e Metabolismo Do GlicogênioDocumento27 páginasGliconeogenese e Metabolismo Do GlicogênioRicardo BorgesAinda não há avaliações
- Gliconeogenese e Via Das Pentoses FosfatosDocumento33 páginasGliconeogenese e Via Das Pentoses Fosfatoscarolaine santos100% (1)
- Metabolismo 2 BioquimicaDocumento13 páginasMetabolismo 2 Bioquimicaalline alvesAinda não há avaliações
- Glicolise e Ciclo de KrebsDocumento12 páginasGlicolise e Ciclo de KrebsLaryssa KlugeAinda não há avaliações
- Base de Dados BQ II - PortuguêsDocumento19 páginasBase de Dados BQ II - PortuguêsZé FerreiraAinda não há avaliações
- GlicóliseDocumento4 páginasGlicóliseGlen daAinda não há avaliações
- BIOQUÍMICA II 02 - Glicólise e GliconeogêneseDocumento8 páginasBIOQUÍMICA II 02 - Glicólise e GliconeogêneseTúlio MaranhãoAinda não há avaliações
- Prova Bioquimica2Documento8 páginasProva Bioquimica2Tatiane MoraisAinda não há avaliações
- Glicolise e GliconeogeneseDocumento49 páginasGlicolise e GliconeogeneseAlline RodriguesAinda não há avaliações
- GlicóliseDocumento4 páginasGlicóliseLivia RossetiAinda não há avaliações
- UntitledDocumento5 páginasUntitledAna RosaAinda não há avaliações
- Carboidratos - MetabolismoDocumento108 páginasCarboidratos - MetabolismoMathews Oviedo100% (1)
- Resumo GlicóliseDocumento6 páginasResumo GlicóliseCamila LopesAinda não há avaliações
- Glicolise e Gliconeogenese Apresentação OkDocumento89 páginasGlicolise e Gliconeogenese Apresentação OkGodronAinda não há avaliações
- GliconeogeneseDocumento4 páginasGliconeogeneseAline PereiraAinda não há avaliações
- Metabolismo Dos CarboidratosDocumento31 páginasMetabolismo Dos CarboidratosRenata AdonesAinda não há avaliações
- Aula II Metabolismo CarboidratosDocumento63 páginasAula II Metabolismo CarboidratosBila BernardesAinda não há avaliações
- Glicogênese, Glicenolise, GliconeogeneseDocumento3 páginasGlicogênese, Glicenolise, GliconeogeneseLaryssa KlugeAinda não há avaliações
- Gliconeogênese, Metabolismo Do Glicogênio e Regulação Da Glicemia PlasmáticaDocumento11 páginasGliconeogênese, Metabolismo Do Glicogênio e Regulação Da Glicemia PlasmáticaFabiana Dantas AlvesAinda não há avaliações
- Bioquimica Metabolica-ResumoDocumento15 páginasBioquimica Metabolica-Resumodani_eleine0% (1)
- Bioquimica - MetabolismoDocumento21 páginasBioquimica - MetabolismoNelma Assis Siqueira100% (1)
- SP 8 - BioquímicaDocumento21 páginasSP 8 - BioquímicaNathally NeriAinda não há avaliações
- GlicoseDocumento8 páginasGlicoselaviAinda não há avaliações
- Metabolismo Do GlicogênioDocumento20 páginasMetabolismo Do Glicogênioamandinha_fc100% (6)
- Trabalho Unidade 2 - Morôni Canella Fraga ScarsiDocumento3 páginasTrabalho Unidade 2 - Morôni Canella Fraga ScarsiMoroni Canella Fraga ScarsiAinda não há avaliações
- Bioenergetica & GlicóliseDocumento32 páginasBioenergetica & Glicólisegiselle rosa ribeiroAinda não há avaliações
- Aula 8-Metab-CarboidratoDocumento28 páginasAula 8-Metab-CarboidratogonsalvessaraivadanielAinda não há avaliações
- Metabolismo Dos GlicídiosDocumento4 páginasMetabolismo Dos GlicídiosMariana BarbosaAinda não há avaliações
- Metabolismo Glicognio Regulao2022Documento35 páginasMetabolismo Glicognio Regulao2022amanda abreuAinda não há avaliações
- Prolactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoNo EverandProlactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoAinda não há avaliações
- Lista Exercícios - Carboidratos e LipídeosDocumento10 páginasLista Exercícios - Carboidratos e LipídeosGuilherme Belak SchmitkeAinda não há avaliações
- 11 e 12 OxidaçãoDocumento1 página11 e 12 OxidaçãoGuilherme Belak SchmitkeAinda não há avaliações
- Lista de Exercícios - EnzimasDocumento5 páginasLista de Exercícios - EnzimasGuilherme Belak SchmitkeAinda não há avaliações
- 13,14 e 15 OxidaçãoDocumento1 página13,14 e 15 OxidaçãoGuilherme Belak SchmitkeAinda não há avaliações
- Atividade Eletro 5Documento2 páginasAtividade Eletro 5Guilherme Belak SchmitkeAinda não há avaliações
- FIBRASDocumento10 páginasFIBRASGuilherme Belak SchmitkeAinda não há avaliações
- Manejo Do LixoDocumento1 páginaManejo Do LixoGuilherme Belak SchmitkeAinda não há avaliações
- FÓRMULAS DRIs PARA CÁLCULO DO GEB E DO GET 24.05.17Documento8 páginasFÓRMULAS DRIs PARA CÁLCULO DO GEB E DO GET 24.05.17ROSANGELA APARECIDA XIMENEZAinda não há avaliações
- Atividade Eletro 3Documento25 páginasAtividade Eletro 3Guilherme Belak SchmitkeAinda não há avaliações
- Quando Os Micro Se MultiplicamDocumento1 páginaQuando Os Micro Se MultiplicamGuilherme Belak SchmitkeAinda não há avaliações
- Plano de Aula Modelos AtomicosDocumento2 páginasPlano de Aula Modelos AtomicosGuilherme Belak SchmitkeAinda não há avaliações
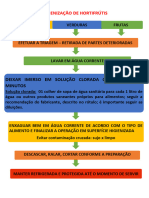
- Fluxograma Higienização de HortifrutisDocumento2 páginasFluxograma Higienização de HortifrutisGuilherme Belak SchmitkeAinda não há avaliações
- Fluxograma Descongelamento de AlimentosDocumento1 páginaFluxograma Descongelamento de AlimentosGuilherme Belak SchmitkeAinda não há avaliações
- Plano de Aula Modelos AtomicosDocumento2 páginasPlano de Aula Modelos AtomicosGuilherme Belak SchmitkeAinda não há avaliações
- Plano de Aula Modelos AtomicosDocumento2 páginasPlano de Aula Modelos AtomicosGuilherme Belak SchmitkeAinda não há avaliações
- Plano de Aula - RadioatividadeDocumento3 páginasPlano de Aula - RadioatividadeGuilherme Belak SchmitkeAinda não há avaliações
- Plano de Aula - Dia Consc. Negra - Misturas e Processos de SeparaçãoDocumento3 páginasPlano de Aula - Dia Consc. Negra - Misturas e Processos de SeparaçãoGuilherme Belak SchmitkeAinda não há avaliações
- Plano de Aula - Dia Consc. Negra - Funções OrgânicasDocumento3 páginasPlano de Aula - Dia Consc. Negra - Funções OrgânicasGuilherme Belak SchmitkeAinda não há avaliações
- AlotropiaPlano de AulaDocumento2 páginasAlotropiaPlano de AulaGuilherme Belak SchmitkeAinda não há avaliações
- M. AAS eDocumento23 páginasM. AAS eHerminio CondulaAinda não há avaliações
- FDR RF6500C 3door DA68-04745F-00 BPTDocumento88 páginasFDR RF6500C 3door DA68-04745F-00 BPTmentecuriosadejimmyAinda não há avaliações
- Avaliação Termoquímica - 2ºano QuímicaDocumento2 páginasAvaliação Termoquímica - 2ºano QuímicaFrannn22duran DuranAinda não há avaliações
- Ficha de Avaliação Fitossanitária - Praça Getúlio Vargas I - LocalizaçãoDocumento150 páginasFicha de Avaliação Fitossanitária - Praça Getúlio Vargas I - LocalizaçãoJamilson da silva juniorAinda não há avaliações
- Reações de Adição EletrofílicaDocumento269 páginasReações de Adição EletrofílicaGuilherme AlmeidaAinda não há avaliações
- Processo de Eletrocoagulação Como Alternativa de Clarificação Do Caldo de Cana Na Produção de Açúcar Cristal - SciELO em Perspectiva - Press ReleasesDocumento5 páginasProcesso de Eletrocoagulação Como Alternativa de Clarificação Do Caldo de Cana Na Produção de Açúcar Cristal - SciELO em Perspectiva - Press ReleasesLetícia Melo JSAinda não há avaliações
- Tabela de Roscas 203-ConexõesDocumento99 páginasTabela de Roscas 203-ConexõesSérgioTeixeiraAraújoAinda não há avaliações
- Materiais de Construção Mecânica 2Documento18 páginasMateriais de Construção Mecânica 2fabiofambhAinda não há avaliações
- FISPQ - Penguard Primer Comp A (Jotun)Documento12 páginasFISPQ - Penguard Primer Comp A (Jotun)zoinhuhcAinda não há avaliações
- WEG Tinta N6.5Documento15 páginasWEG Tinta N6.5Jeovane VasconcelosAinda não há avaliações
- Questionário Abendi LPDocumento15 páginasQuestionário Abendi LPjonathan wesley da costa100% (3)
- Regulacao Do Equilibrio Hidrico e Acido BaseDocumento19 páginasRegulacao Do Equilibrio Hidrico e Acido BaseMario PontavidaAinda não há avaliações
- Lista de Exercícos de GasesDocumento5 páginasLista de Exercícos de GasesdanielmirandatmAinda não há avaliações
- Relatorio FisicaDocumento9 páginasRelatorio FisicaJf FernandesAinda não há avaliações
- Guia Prático DiamaxDocumento24 páginasGuia Prático DiamaxHigor RibeiroAinda não há avaliações
- Composiçao EstequimetricaDocumento20 páginasComposiçao EstequimetricaJaime ChogaAinda não há avaliações
- Névoa Salina RelatórioDocumento4 páginasNévoa Salina Relatórioyuri maximoAinda não há avaliações
- Fispq-Smart B-226Documento11 páginasFispq-Smart B-226João Vitor RagonhaAinda não há avaliações
- Central de Material e Esterilização Aula 2Documento34 páginasCentral de Material e Esterilização Aula 2Adlesio jose0% (1)
- Farmacocinéctica - Aula 1Documento35 páginasFarmacocinéctica - Aula 1Catarina CuryAinda não há avaliações
- Materia CorpuscularDocumento9 páginasMateria CorpuscularEvaristo Paulo CassomaAinda não há avaliações
- Exercicios 2 e 3Documento5 páginasExercicios 2 e 3Eng. Luiz Mario J. Borges100% (1)
- Exercicios IsomeriaDocumento9 páginasExercicios IsomeriaVinicius MoreiraAinda não há avaliações
- FembendazolDocumento1 páginaFembendazolcelmorcelliAinda não há avaliações
- Relatório BromatologiaDocumento11 páginasRelatório BromatologialuanavanAinda não há avaliações
- ALPHAWORLD - IndividualidadesDocumento37 páginasALPHAWORLD - IndividualidadesJunior SantosAinda não há avaliações
- Templete - Analítica - Qualitativa - Template 2Documento3 páginasTemplete - Analítica - Qualitativa - Template 2Jessiane QueirozAinda não há avaliações
- Catalogo - Calhas e LeitosDocumento16 páginasCatalogo - Calhas e LeitosLuiz Sérgio SilvaAinda não há avaliações
- Apostila - Projeto Hidráulico BásicoDocumento58 páginasApostila - Projeto Hidráulico BásicoLuiz Gabriel Belling100% (1)