Você também pode gostar
- Slides Perfil Sensorial 2Documento136 páginasSlides Perfil Sensorial 2Karla Niana Monteiro Rezende100% (7)
- AULA 3 Transformação Músculo em CarneDocumento47 páginasAULA 3 Transformação Músculo em CarneAgro PetAinda não há avaliações
- Medcel Ginecologia - Vol 3Documento160 páginasMedcel Ginecologia - Vol 3Karoline CordeiroAinda não há avaliações
- Swallowing Water - Siggy ShadeDocumento143 páginasSwallowing Water - Siggy ShadeMaysaS AmorimAinda não há avaliações
- Fisiologia Musculo EsqueleticoDocumento14 páginasFisiologia Musculo Esqueleticojoice alvesAinda não há avaliações
- ProteínaDocumento23 páginasProteínaTyagoB BustorfAinda não há avaliações
- Ap - Uso de Bioestimulantes Na Indução de Resistência para o Manejo FitossanitárioDocumento23 páginasAp - Uso de Bioestimulantes Na Indução de Resistência para o Manejo FitossanitárioAnna Carolina BarbozaAinda não há avaliações
- Apostila de Patologia Geral 2021 (Aires)Documento70 páginasApostila de Patologia Geral 2021 (Aires)Vitória FernandesAinda não há avaliações
- Aula 9 Degradacao Oxidativa de ProteinasDocumento29 páginasAula 9 Degradacao Oxidativa de ProteinasA Lendária Eguinha PocotóAinda não há avaliações
- Resumo - Integração MetabólicaDocumento18 páginasResumo - Integração MetabólicamuradihachichuanAinda não há avaliações
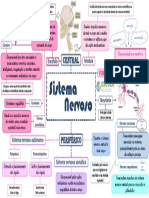
- Mapa Mental Sistema NervosoDocumento1 páginaMapa Mental Sistema NervosoVanessa MoreiraAinda não há avaliações
- Metabolismo Dos AminoácidosDocumento7 páginasMetabolismo Dos AminoácidosErica AdrianoAinda não há avaliações
- AMINOÁCIDOSDocumento19 páginasAMINOÁCIDOSRodolfo CrespoAinda não há avaliações
- Bioqumicaii09 Ciclodauriaarlindonetto 120627022145 Phpapp02Documento0 páginaBioqumicaii09 Ciclodauriaarlindonetto 120627022145 Phpapp02Vivian Leão VeterináriaAinda não há avaliações
- Os Aminoácidos Não São Armazenados Pelo CorpoDocumento6 páginasOs Aminoácidos Não São Armazenados Pelo CorpoadrianokotAinda não há avaliações
- Nitrogen I o Raquel MDocumento13 páginasNitrogen I o Raquel MgeizeAinda não há avaliações
- Metabolismo de ProteínasDocumento26 páginasMetabolismo de ProteínasGuilherme ZulimAinda não há avaliações
- Metabolismo de ProteinasDocumento2 páginasMetabolismo de ProteinasMirelleTavaresAinda não há avaliações
- Thumb - Metabolismo Do NitrogênioDocumento17 páginasThumb - Metabolismo Do NitrogênioflaviaAinda não há avaliações
- Metabolismo ProteínasDocumento6 páginasMetabolismo ProteínasSarah GabriellaAinda não há avaliações
- (Resumo) Bioquim - Catabolismo de ProteinasDocumento16 páginas(Resumo) Bioquim - Catabolismo de Proteinasabresser100% (2)
- Proteinas Atividade FisicaDocumento7 páginasProteinas Atividade FisicaKarime MendonçaAinda não há avaliações
- Bioquímica - Catabolismo de AminoácidosDocumento7 páginasBioquímica - Catabolismo de AminoácidosMarcela Lima de BritoAinda não há avaliações
- Metabolismo de AminoácidosDocumento8 páginasMetabolismo de AminoácidosDenisar PalmitoAinda não há avaliações
- 6 - Oxidação de Aminoácidos e Ciclo Da Ureia ResumoDocumento7 páginas6 - Oxidação de Aminoácidos e Ciclo Da Ureia ResumoCaruma GuerreiroAinda não há avaliações
- Metabolismo de ProteínasDocumento12 páginasMetabolismo de ProteínasMonica AlexandreAinda não há avaliações
- Oxidação Dos Aminoácidos e Produção de UréiaDocumento20 páginasOxidação Dos Aminoácidos e Produção de UréiaHenrique BarbosaAinda não há avaliações
- BQ - Metabolismo de AminoácidosDocumento8 páginasBQ - Metabolismo de AminoácidosYuri RochaAinda não há avaliações
- Roteiro - Metabolismo de Aminoácidos e ProteínasDocumento4 páginasRoteiro - Metabolismo de Aminoácidos e ProteínasMarina StrossiAinda não há avaliações
- Ciclo Da UreiaDocumento65 páginasCiclo Da Ureiamellogenilza88Ainda não há avaliações
- Aula 7 - Degradação Dos Alfa-CetoácidosDocumento5 páginasAula 7 - Degradação Dos Alfa-CetoácidosArielle MoraesAinda não há avaliações
- Proteínas e Enzimas Iberê Catolica-1Documento42 páginasProteínas e Enzimas Iberê Catolica-1Anderson R. de OliveiraAinda não há avaliações
- Metabolismo de Proteínas PDFDocumento15 páginasMetabolismo de Proteínas PDFJandderson CostaAinda não há avaliações
- Revisão para BioquímicaDocumento4 páginasRevisão para BioquímicaSthephanne PaolaAinda não há avaliações
- Transaminação de AADocumento7 páginasTransaminação de AABeatriz Souza LimaAinda não há avaliações
- Slides MetabolismoDocumento22 páginasSlides MetabolismofierymAinda não há avaliações
- Integração MetabolicaDocumento17 páginasIntegração MetabolicaDjinane AndreaAinda não há avaliações
- Bioquímica: Aula 4Documento16 páginasBioquímica: Aula 4Tuani de MirandaAinda não há avaliações
- 3 - Proteína e EnzimasDocumento8 páginas3 - Proteína e EnzimasIsabela Zucoloto MasoliniAinda não há avaliações
- 1química e Metabolismo de Aminoácidos - MaxDocumento42 páginas1química e Metabolismo de Aminoácidos - MaxhhfzpmbvhsAinda não há avaliações
- Metabolismo ProteinasDocumento33 páginasMetabolismo ProteinastaefrouAinda não há avaliações
- Bioquimica 10-20Documento11 páginasBioquimica 10-20Isabelle SousaAinda não há avaliações
- Material Aula EletroforeseDocumento45 páginasMaterial Aula EletroforeseKarol SilvaAinda não há avaliações
- Aula 09 - Metabolismo Das Proteínas No Exercício FísicoDocumento4 páginasAula 09 - Metabolismo Das Proteínas No Exercício FísicoIgor ChicoletAinda não há avaliações
- Fichamento Sobre Proteínas - MetabolismoDocumento6 páginasFichamento Sobre Proteínas - MetabolismoAline da Silva Fernandes MatiasAinda não há avaliações
- Bioquimica Metabolismo ProteinasDocumento16 páginasBioquimica Metabolismo ProteinasNoelen FrancoAinda não há avaliações
- Metabolismo Do NitrogênioDocumento14 páginasMetabolismo Do NitrogênioVandik CandidoAinda não há avaliações
- MacronutrientesDocumento15 páginasMacronutrientesLohraine FreitasAinda não há avaliações
- Fechamento P4 UC IVDocumento13 páginasFechamento P4 UC IVLuis Mateus Camelo França MartinsAinda não há avaliações
- Degradação e Síntese Dos AminiácidosDocumento3 páginasDegradação e Síntese Dos AminiácidosJordana MesquitaAinda não há avaliações
- Sessão 7Documento5 páginasSessão 7Gabriel PiresAinda não há avaliações
- Metabolismo Dos AminoacidosDocumento17 páginasMetabolismo Dos AminoacidosRodrigoPânicoAinda não há avaliações
- PROTEINASDocumento7 páginasPROTEINASlourran19Ainda não há avaliações
- Aula 6 - Metabolismo Dos AminoácidosDocumento6 páginasAula 6 - Metabolismo Dos AminoácidosArielle MoraesAinda não há avaliações
- Biossintese de AminoacidosDocumento13 páginasBiossintese de AminoacidosJuliane Ketlei0% (1)
- Roteiro Elisa Aula 6 - Metabolismo de Aminoácidos e Ciclo Da UréiaDocumento8 páginasRoteiro Elisa Aula 6 - Metabolismo de Aminoácidos e Ciclo Da UréiaElisa de AlmeidaAinda não há avaliações
- 1253 Fisiologia+do+exercicioDocumento50 páginas1253 Fisiologia+do+exercicioDaniel Matos SilvaAinda não há avaliações
- Unidade II - Tema 3 - ProteínasDocumento15 páginasUnidade II - Tema 3 - ProteínasINSTITUTO DE ENSINO INESPECAinda não há avaliações
- Oxidação de Aminoácidos e Produção de UreiaDocumento10 páginasOxidação de Aminoácidos e Produção de UreiaErika BennaiaAinda não há avaliações
- Ciclo Da Ureia1Documento6 páginasCiclo Da Ureia1HENRIQUEAinda não há avaliações
- Metabolismo Dos Aminoácidos MonografiaDocumento3 páginasMetabolismo Dos Aminoácidos MonografiaDianaSantosAinda não há avaliações
- Processos CitoplasmáticosDocumento8 páginasProcessos CitoplasmáticosLone GhostAinda não há avaliações
- Macromoleculas e Respiração CelularDocumento7 páginasMacromoleculas e Respiração CelularGabriela OliveiraAinda não há avaliações
- Metabolismo Dos GlicídiosDocumento4 páginasMetabolismo Dos GlicídiosMariana BarbosaAinda não há avaliações
- PDF 20230228 221028 0000Documento29 páginasPDF 20230228 221028 0000Leonardo Wanderley FilhoAinda não há avaliações
- 7670-Texto Do Artigo-28879-1-10-20091006Documento13 páginas7670-Texto Do Artigo-28879-1-10-20091006Mariana BarbosaAinda não há avaliações
- Pseudomelanose e Embebição Por HemoglobinaDocumento10 páginasPseudomelanose e Embebição Por HemoglobinaMariana BarbosaAinda não há avaliações
- EquinosDocumento8 páginasEquinosMariana BarbosaAinda não há avaliações
- Farelo de Urucum em PoedeirasDocumento3 páginasFarelo de Urucum em PoedeirasMariana BarbosaAinda não há avaliações
- Patologia Geral 2015 A - Aula 1Documento11 páginasPatologia Geral 2015 A - Aula 1Mariana BarbosaAinda não há avaliações
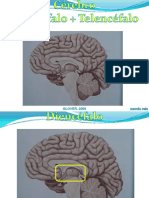
- NRT 9 CérebroDocumento59 páginasNRT 9 CérebroMariana BarbosaAinda não há avaliações
- Conteudo Prog Nutricao AnimalDocumento1 páginaConteudo Prog Nutricao AnimalMariana BarbosaAinda não há avaliações
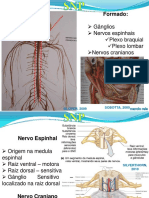
- NRT 5 Medula EspinhalDocumento22 páginasNRT 5 Medula EspinhalMariana BarbosaAinda não há avaliações
- Prática 11 - Reações de Identificação de GlicídiosDocumento1 páginaPrática 11 - Reações de Identificação de GlicídiosMariana BarbosaAinda não há avaliações
- NRT 7 SNPDocumento20 páginasNRT 7 SNPMariana BarbosaAinda não há avaliações
- NRT 6 Tronco EncefálicoDocumento17 páginasNRT 6 Tronco EncefálicoMariana BarbosaAinda não há avaliações
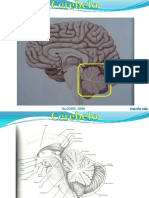
- NRT 8 CerebeloDocumento12 páginasNRT 8 CerebeloMariana BarbosaAinda não há avaliações
- NRT 9 CérebroDocumento59 páginasNRT 9 CérebroMariana BarbosaAinda não há avaliações
- NRT 3 SNADocumento14 páginasNRT 3 SNAMariana BarbosaAinda não há avaliações
- NRT 5 Medula EspinhalDocumento22 páginasNRT 5 Medula EspinhalMariana BarbosaAinda não há avaliações
- NRT 1 IntroduçãoDocumento24 páginasNRT 1 IntroduçãoMariana BarbosaAinda não há avaliações
- NRT 2 Meninge VascularizaçãoDocumento31 páginasNRT 2 Meninge VascularizaçãoMariana BarbosaAinda não há avaliações
- Prática 14 - Identificação Dos Componentes Do LeiteDocumento2 páginasPrática 14 - Identificação Dos Componentes Do LeiteMariana BarbosaAinda não há avaliações
- NF Ranger HNP7781Documento1 páginaNF Ranger HNP7781Mariana BarbosaAinda não há avaliações
- Prática 10 - Extração e Caracterização de Glicídios Na BatataDocumento1 páginaPrática 10 - Extração e Caracterização de Glicídios Na BatataMariana BarbosaAinda não há avaliações
- Prática 13 - Reaçãode de Identificação Dos LipídiosDocumento2 páginasPrática 13 - Reaçãode de Identificação Dos LipídiosMariana BarbosaAinda não há avaliações
- 5° Workshop de Análise Corporal v4 PDFDocumento34 páginas5° Workshop de Análise Corporal v4 PDFJefAinda não há avaliações
- Prática 12 - Preparo e Aplicação Do Reativo de Benedict - Glicose Na UrinaDocumento2 páginasPrática 12 - Preparo e Aplicação Do Reativo de Benedict - Glicose Na UrinaMariana BarbosaAinda não há avaliações
- Prática 7 - Determinação de Albumina Na UrinaDocumento1 páginaPrática 7 - Determinação de Albumina Na UrinaMariana BarbosaAinda não há avaliações
- Prática 11 - Reações de Identificação de GlicídiosDocumento1 páginaPrática 11 - Reações de Identificação de GlicídiosMariana BarbosaAinda não há avaliações
- Prática 5 - Reativo de BiuretoDocumento1 páginaPrática 5 - Reativo de BiuretoMariana BarbosaAinda não há avaliações
- Prática 9 - Atividade FermentativaDocumento2 páginasPrática 9 - Atividade FermentativaMariana BarbosaAinda não há avaliações
- Prática 8 - Atividade EnzimáticaDocumento2 páginasPrática 8 - Atividade EnzimáticaMariana BarbosaAinda não há avaliações
- Prática 6 - Precipitação de ProteínasDocumento1 páginaPrática 6 - Precipitação de ProteínasMariana BarbosaAinda não há avaliações
- Prática 11 - Reações de Identificação de GlicídiosDocumento1 páginaPrática 11 - Reações de Identificação de GlicídiosMariana BarbosaAinda não há avaliações
- Antigenos e Anticorpos PDFDocumento2 páginasAntigenos e Anticorpos PDFMonique MeirellesAinda não há avaliações
- Novo Guia Do Aumento PenianoDocumento14 páginasNovo Guia Do Aumento PenianoDicas para GestanteAinda não há avaliações
- Prática 8 - Atividade EnzimáticaDocumento2 páginasPrática 8 - Atividade EnzimáticaMariana BarbosaAinda não há avaliações
- E-Book - Amamentação DescomplicadaDocumento27 páginasE-Book - Amamentação Descomplicadaalex silvaAinda não há avaliações
- Leucócitos: Mielócitos: Metamielócitos: Bastões ( ) : Segmentados: Linfócitos: Monócitos: Eosinófilos: BasófilosDocumento8 páginasLeucócitos: Mielócitos: Metamielócitos: Bastões ( ) : Segmentados: Linfócitos: Monócitos: Eosinófilos: BasófilosThamara FerreiraAinda não há avaliações
- NEUROTRANSMISSORESDocumento2 páginasNEUROTRANSMISSORESMaria Eduarda BitúAinda não há avaliações
- Alteracoes Do IntersticioDocumento34 páginasAlteracoes Do IntersticioPatrícia S. SantosAinda não há avaliações
- Prova II Diversidade de Fanerógamos UniasselviDocumento6 páginasProva II Diversidade de Fanerógamos UniasselviMauricio CamposAinda não há avaliações
- Do Inconsciente Vital À Consciência Cósmica. Maria Carmen Souza DallalanaDocumento70 páginasDo Inconsciente Vital À Consciência Cósmica. Maria Carmen Souza DallalanaRita BellocchioAinda não há avaliações
- Aula5 TECIDO CONJUTIVO Parte2Documento26 páginasAula5 TECIDO CONJUTIVO Parte2Zakeu QuellyAinda não há avaliações
- Neoplasias - PatologiaDocumento2 páginasNeoplasias - PatologiaKésia SilvaAinda não há avaliações
- 10 Sistema EndocrinoDocumento25 páginas10 Sistema EndocrinoGlaucia BentesAinda não há avaliações
- Album Didatico de Anatomia VegetalDocumento65 páginasAlbum Didatico de Anatomia VegetalJose Franco NaviaAinda não há avaliações
- BIBLIA DE BIOLOGIA Comecando Com GlicideosDocumento15 páginasBIBLIA DE BIOLOGIA Comecando Com GlicideosviniciusAinda não há avaliações
- Entendendo o HemogramaDocumento10 páginasEntendendo o HemogramaAngela GontijoAinda não há avaliações
- Tema - Células e Organelas - Habilidades - Identificar Os Elementos Básicos Que Compõem A Célula, Bem Como Suas Funções.Documento7 páginasTema - Células e Organelas - Habilidades - Identificar Os Elementos Básicos Que Compõem A Célula, Bem Como Suas Funções.ANA ANGELICA CORDEIRO DE SOUSAAinda não há avaliações
- LipídeosDocumento121 páginasLipídeosGILBERTH SILVA NUNESAinda não há avaliações
- Atividade de Ciências - 4º Ano (14-10) - ClasseDocumento4 páginasAtividade de Ciências - 4º Ano (14-10) - ClasseIeda CerqueiraAinda não há avaliações
- Estudo Dirigido - Glicídios e Metabolismo Dos GlicídiosDocumento2 páginasEstudo Dirigido - Glicídios e Metabolismo Dos GlicídiosMariana BarbosaAinda não há avaliações
- 1461703808103tabela Exames Laboratoriais 2016Documento10 páginas1461703808103tabela Exames Laboratoriais 2016Vanessa NatalAinda não há avaliações
- Ap Osti Lad e Patologia G Eral: @dentologaDocumento81 páginasAp Osti Lad e Patologia G Eral: @dentologaHevellyn SouzaAinda não há avaliações
- ANATO - Aula 1 ReprodutorDocumento10 páginasANATO - Aula 1 ReprodutorRoiter BarcelosAinda não há avaliações