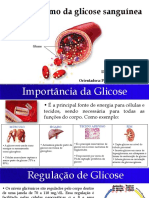
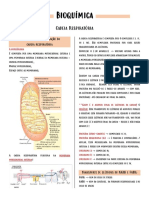
Você também pode gostar
- Estudo Dirigido BioquimicaDocumento5 páginasEstudo Dirigido Bioquimicaadelsonribeiro100% (4)
- Bioquimica Aplicada A SaudeDocumento9 páginasBioquimica Aplicada A SaudeNaiara OliveiraAinda não há avaliações
- Cadeia Transportadora de ElétronsDocumento1 páginaCadeia Transportadora de ElétronsElisa CamposAinda não há avaliações
- CiclosDocumento62 páginasCiclosMJTAinda não há avaliações
- Pâncreas - Apostilas - Fisiologia PDFDocumento6 páginasPâncreas - Apostilas - Fisiologia PDFMatheus CostaAinda não há avaliações
- Metabolismo Do GlicogenioDocumento3 páginasMetabolismo Do GlicogenioAlice AlencarAinda não há avaliações
- Diabetes MellitusDocumento16 páginasDiabetes Mellitusana paula de castro costaAinda não há avaliações
- Metabolismo Cho PTN LipDocumento20 páginasMetabolismo Cho PTN LipMariana CarvalhoAinda não há avaliações
- Aula 6 - PâncreasDocumento2 páginasAula 6 - Pâncreasraquel luzAinda não há avaliações
- FT 512 Exercicios CarboidratosDocumento3 páginasFT 512 Exercicios CarboidratosAna Julia Da MataAinda não há avaliações
- Sistema Digestório - Bioquímica - Passei DiretoDocumento5 páginasSistema Digestório - Bioquímica - Passei DiretoRenata MendesAinda não há avaliações
- Continuação: Carboidratos + Sua Digestão e AbsorçãoDocumento8 páginasContinuação: Carboidratos + Sua Digestão e AbsorçãoAna VitóriaAinda não há avaliações
- Aula 9 - Glicólise e Destinos Do PiruvatoDocumento53 páginasAula 9 - Glicólise e Destinos Do PiruvatoFernando BritoAinda não há avaliações
- Diabetes Mellitus TeoricaDocumento12 páginasDiabetes Mellitus TeoricaThais OliveiraAinda não há avaliações
- Glicogenólise - BioquímicaDocumento3 páginasGlicogenólise - BioquímicaDeywison Dábilo - MEDICINAAinda não há avaliações
- Metabolismo CarboidratosDocumento14 páginasMetabolismo CarboidratosIsadora ScopelAinda não há avaliações
- Transportadores de Glicose Tecidos Dependentes e Independentes de InsulinaDocumento6 páginasTransportadores de Glicose Tecidos Dependentes e Independentes de InsulinaCamila100% (1)
- Metabolismo de CHO (Questões)Documento10 páginasMetabolismo de CHO (Questões)Iana Ferreira100% (1)
- Metabolismo de Carboidratos - Glicólise e Formação de Acetil-CoAAmido - Lactose e SacaroseDocumento22 páginasMetabolismo de Carboidratos - Glicólise e Formação de Acetil-CoAAmido - Lactose e SacaroseSAngela MarquesAinda não há avaliações
- Gliconeogênese Seminario Rodolfo 29 de Outubro 2021.2Documento7 páginasGliconeogênese Seminario Rodolfo 29 de Outubro 2021.2MC CHACALAinda não há avaliações
- Resumo - Endocrinologia - Fabiana Getulino ATM2023Documento6 páginasResumo - Endocrinologia - Fabiana Getulino ATM2023TaironeAinda não há avaliações
- 15) NeoglicogêneseDocumento4 páginas15) NeoglicogêneseMary FreitasAinda não há avaliações
- Metabolismo Dos GlicídiosDocumento4 páginasMetabolismo Dos GlicídiosMariana BarbosaAinda não há avaliações
- P2 - ArmazenamentoDocumento9 páginasP2 - ArmazenamentoPedro Henrique Almeida de BritoAinda não há avaliações
- Estrategias Regulacão Período Pós-Absortivo e AbsortivoDocumento24 páginasEstrategias Regulacão Período Pós-Absortivo e AbsortivoJanaina Cunha Barbosa DalloAinda não há avaliações
- Tut VDocumento5 páginasTut VGabriel PiresAinda não há avaliações
- Gli Nóli: OBS: Se A Gordura É Mais Eficiente, Energeticamente, Por Que Ela Não É Usada No Lugar Do Glicogênio?Documento3 páginasGli Nóli: OBS: Se A Gordura É Mais Eficiente, Energeticamente, Por Que Ela Não É Usada No Lugar Do Glicogênio?Deywison Dábilo - MEDICINAAinda não há avaliações
- Introdução Ao Metabolismo EnergéticoDocumento2 páginasIntrodução Ao Metabolismo EnergéticoDavi ÁlefeAinda não há avaliações
- GlicoseDocumento8 páginasGlicoselaviAinda não há avaliações
- Pancreas e AdrenalDocumento11 páginasPancreas e AdrenalDarlanSilvaAinda não há avaliações
- Bioquímica - Ciclo Alimentado e JejumDocumento9 páginasBioquímica - Ciclo Alimentado e JejumPedro CintraAinda não há avaliações
- Fichamento Sobre CarboidratosDocumento16 páginasFichamento Sobre CarboidratosPatricia AlvesAinda não há avaliações
- Via Glicolítica IVDocumento28 páginasVia Glicolítica IVSabrina AraujoAinda não há avaliações
- Trabalho GlicogenóliseDocumento6 páginasTrabalho GlicogenóliseJulia MoraesAinda não há avaliações
- Bioquimica 03 Carboidratos 1Documento20 páginasBioquimica 03 Carboidratos 1Marllon MarinsAinda não há avaliações
- Aula 4 - LPF - Diabetes Gestacional - Teste de Tolerância Oral ÀDocumento11 páginasAula 4 - LPF - Diabetes Gestacional - Teste de Tolerância Oral Àgabriella.ferreira.inacioAinda não há avaliações
- Energia Da Urina (EU)Documento10 páginasEnergia Da Urina (EU)j.efersonsanttanaaAinda não há avaliações
- Questionário Glicólise e Catabolismo Das HexosesDocumento6 páginasQuestionário Glicólise e Catabolismo Das HexosesRaffael Batista MarquesAinda não há avaliações
- Farmacologia Da DiabetesDocumento10 páginasFarmacologia Da DiabetesAfiliado Lendário100% (1)
- Diabetes Mellitus e LipidogrmaDocumento26 páginasDiabetes Mellitus e LipidogrmaJessica MoraesAinda não há avaliações
- Glicogênese, Glicenolise, GliconeogeneseDocumento3 páginasGlicogênese, Glicenolise, GliconeogeneseLaryssa KlugeAinda não há avaliações
- Slide Sobre: Metabolismo Da GlicoseDocumento18 páginasSlide Sobre: Metabolismo Da Glicosedebora machado fonseca100% (1)
- Biossintese de Carboidratos BioquímicaDocumento6 páginasBiossintese de Carboidratos BioquímicaMarcela Lima de BritoAinda não há avaliações
- Curva GlicêmicaDocumento4 páginasCurva GlicêmicaRê SilvaAinda não há avaliações
- Monografia Diabetes 01Documento16 páginasMonografia Diabetes 01italofg100% (1)
- BioquimiscDocumento7 páginasBioquimiscJuca Do sabugueiro loucoAinda não há avaliações
- Metabolismo Dos CarboidratosDocumento21 páginasMetabolismo Dos CarboidratosRamon BrazAinda não há avaliações
- Regulação Do Metabolismo - HORMÔNIOS 2023Documento42 páginasRegulação Do Metabolismo - HORMÔNIOS 2023Nádia SousaAinda não há avaliações
- Bases de Nutrição: CarboidratosDocumento8 páginasBases de Nutrição: CarboidratosAna Beatriz FreitasAinda não há avaliações
- Problema 01 10Documento23 páginasProblema 01 10Karoline Silva ZampronhoAinda não há avaliações
- Metabolismo Dos Carboidratos, Lipã Dios e Proteã NasDocumento8 páginasMetabolismo Dos Carboidratos, Lipã Dios e Proteã NasCynthia MunizAinda não há avaliações
- Integração Metabólica e OligoelementosDocumento2 páginasIntegração Metabólica e OligoelementosPATRICIA AMORIMAinda não há avaliações
- Uma Panorâmica Geral Das Vias MetabólicasDocumento4 páginasUma Panorâmica Geral Das Vias MetabólicasVanessa TomazAinda não há avaliações
- Bioquímica VeterináriaDocumento7 páginasBioquímica Veterináriajéssica catarinaAinda não há avaliações
- Glicolise e Gliconeogenese Apresentação OkDocumento89 páginasGlicolise e Gliconeogenese Apresentação OkGodronAinda não há avaliações
- Disturbio Do Metabolismo GlicídicoDocumento21 páginasDisturbio Do Metabolismo GlicídicoJefferson LizAinda não há avaliações
- Resumo P2 BioquímicaDocumento21 páginasResumo P2 BioquímicaAnna Luíza AugustoAinda não há avaliações
- Metabolismo Dos CarboidratosDocumento9 páginasMetabolismo Dos CarboidratosRodrigo AcioliAinda não há avaliações
- BIOQUÍMICA II 02 - Glicólise e Gliconeogênese PDFDocumento8 páginasBIOQUÍMICA II 02 - Glicólise e Gliconeogênese PDFCharles Lameira ValenteAinda não há avaliações
- Aula 5Documento9 páginasAula 5Afonso Henriques NunesAinda não há avaliações
- Turmadefevereiro-Biologia1-Respiração Celular-03-05-2023Documento11 páginasTurmadefevereiro-Biologia1-Respiração Celular-03-05-2023Gabriela de Sousa Leal ReisAinda não há avaliações
- Questões Pós Aula Resolvidas - Trabalho Prático 3Documento3 páginasQuestões Pós Aula Resolvidas - Trabalho Prático 3Lucia ElvasAinda não há avaliações
- Aula Ciclo Krebs e Cadeia de Transporte de ElétronsDocumento32 páginasAula Ciclo Krebs e Cadeia de Transporte de ElétronsRafael TorreAinda não há avaliações
- Artigo 2Documento2 páginasArtigo 2Nathália Dias NascimentoAinda não há avaliações
- Cadeia RespiratóriaDocumento4 páginasCadeia RespiratóriaAndré Duarte CanellasAinda não há avaliações
- Slides Aula Cadeia RespiratóriaDocumento5 páginasSlides Aula Cadeia Respiratóriaalicercio cajamuraAinda não há avaliações
- Aula 7 MitocondriasDocumento14 páginasAula 7 MitocondriasAna Marcia CarvalhoAinda não há avaliações
- Anotações BioquímicaDocumento10 páginasAnotações BioquímicaAnderson Souza de AssisAinda não há avaliações
- Membrana Interna Da Mitocôndria: Tutoria de IM 2021/1 Vivianny Vitória Valverde RamosDocumento2 páginasMembrana Interna Da Mitocôndria: Tutoria de IM 2021/1 Vivianny Vitória Valverde RamosEllochAinda não há avaliações
- 12° ResumoDocumento5 páginas12° ResumoRafaela de Jesus OliveiraAinda não há avaliações
- MT MaxSolve AbrDocumento13 páginasMT MaxSolve AbrVitor Ribeiro100% (1)
- Aula 09 - Fosforilação OxidativaDocumento7 páginasAula 09 - Fosforilação OxidativaMarcela Lima de BritoAinda não há avaliações
- Slides Aula 11. CTE e Fosforilação OxidativaDocumento26 páginasSlides Aula 11. CTE e Fosforilação OxidativaÂngelo AbnerAinda não há avaliações
- A Cadeia de Transporte de ElétronsDocumento8 páginasA Cadeia de Transporte de ElétronsSamilly BarbosaAinda não há avaliações
- Fosforilação OxidativaDocumento31 páginasFosforilação Oxidativatitalves100% (1)
- 9133-Texto Do Artigo - 49250-2-10-20201022Documento13 páginas9133-Texto Do Artigo - 49250-2-10-20201022Ronildo CafféAinda não há avaliações
- Seminario de Bioquimica OfcDocumento57 páginasSeminario de Bioquimica OfcDébora CarvalhoAinda não há avaliações
- 8 - TP8 - FraccionamentoDocumento6 páginas8 - TP8 - FraccionamentoDiogo OliveiraAinda não há avaliações
- Visão Geral Da Fosforilação OxidativaDocumento6 páginasVisão Geral Da Fosforilação OxidativaAna Paula AzevedoAinda não há avaliações
- Aulas Teóricas Graduação Fisiologia VegetalDocumento102 páginasAulas Teóricas Graduação Fisiologia VegetalVALDINEI JUNIO BRITO VILELAAinda não há avaliações
- NAD+ No Envelhecimento Mecanismos Moleculares eDocumento5 páginasNAD+ No Envelhecimento Mecanismos Moleculares eRodrigo AlvesAinda não há avaliações
- MONITORIA BASES - Caderno de Questões 2022.2Documento20 páginasMONITORIA BASES - Caderno de Questões 2022.2Dominique SakagamiAinda não há avaliações
- Questões Integração MetabólicaDocumento7 páginasQuestões Integração MetabólicaJoao LucasAinda não há avaliações
- Slides Cadeia Transportadora de EletronsDocumento14 páginasSlides Cadeia Transportadora de EletronscelsogdjAinda não há avaliações