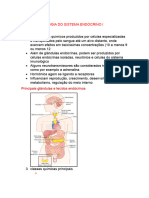
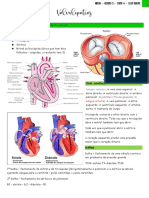
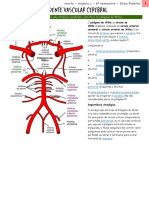
Você também pode gostar
- Protocolo Sinatra CardiopatiaDocumento3 páginasProtocolo Sinatra Cardiopatiasfleandro_67Ainda não há avaliações
- Síndromes pleurais e pulmonares: derrame pleural, pneumotórax e causasDocumento14 páginasSíndromes pleurais e pulmonares: derrame pleural, pneumotórax e causaselisaAinda não há avaliações
- O Ciclo de KrebsDocumento2 páginasO Ciclo de KrebsCamila OliveiraAinda não há avaliações
- Regulação do comportamento alimentarDocumento53 páginasRegulação do comportamento alimentarKarina Costa PaesAinda não há avaliações
- BioquímicaDocumento220 páginasBioquímicaJoão Matheus100% (1)
- Dimensões do movimentoDocumento16 páginasDimensões do movimentoLuís DiasAinda não há avaliações
- Resumo - Ciclo de KrebsDocumento4 páginasResumo - Ciclo de KrebsLAIS VERONEZ MARQUES BARROSAinda não há avaliações
- 3800 Questões de Biologia Com Gabarito PDFDocumento526 páginas3800 Questões de Biologia Com Gabarito PDFKaylane50% (2)
- Doenças Coronarianas - ResumoDocumento30 páginasDoenças Coronarianas - ResumoelisaAinda não há avaliações
- Exercícios Respiração Celular AeróbiaDocumento5 páginasExercícios Respiração Celular AeróbiaAline ChristineAinda não há avaliações
- Fisiopatologia dos principais distúrbios endócrinosDocumento150 páginasFisiopatologia dos principais distúrbios endócrinosRosângela Vasconcelos100% (2)
- Manual de Ergonomia - Adaptando o Trabalho Ao Homem - Etienne GrandjeanDocumento174 páginasManual de Ergonomia - Adaptando o Trabalho Ao Homem - Etienne GrandjeanRafAinda não há avaliações
- Ascite: causas, sinais e diagnósticoDocumento4 páginasAscite: causas, sinais e diagnósticoelisa100% (1)
- 2008 11 14 11 18 28 390 - Testes - AvaliacaoDocumento18 páginas2008 11 14 11 18 28 390 - Testes - AvaliacaoJoana CunhaAinda não há avaliações
- ArtigoDocumento52 páginasArtigoAluana Santana CarlosAinda não há avaliações
- BIOQUIMICA - AULA 9 - Integração MetabólicaDocumento21 páginasBIOQUIMICA - AULA 9 - Integração MetabólicaLP BarbosaAinda não há avaliações
- 258-Texto Do Artigo-656-1-10-20070523 PDFDocumento15 páginas258-Texto Do Artigo-656-1-10-20070523 PDFRaissa GontijoAinda não há avaliações
- ENTENDER O METABOLISMO DA GLICOSE PELO PÂNCREASDocumento5 páginasENTENDER O METABOLISMO DA GLICOSE PELO PÂNCREASBianca AmaralAinda não há avaliações
- Função Gastrointestinal emDocumento3 páginasFunção Gastrointestinal emGustavo AbecassisAinda não há avaliações
- Funções do Pâncreas emDocumento5 páginasFunções do Pâncreas emMiguel MPAinda não há avaliações
- Funções do Pâncreas emDocumento5 páginasFunções do Pâncreas emMiguel MPAinda não há avaliações
- Diabetes: Soi Iv - Apg 03Documento8 páginasDiabetes: Soi Iv - Apg 03Melissa TolentinoAinda não há avaliações
- Integração MetabólicaDocumento35 páginasIntegração MetabólicaRicardo BorgesAinda não há avaliações
- Fisiologia do pâncreasDocumento2 páginasFisiologia do pâncreasraquel luzAinda não há avaliações
- Roteiro Do Laboratório Morfofuncional: Curso de MedicinaDocumento14 páginasRoteiro Do Laboratório Morfofuncional: Curso de MedicinabluboarettoAinda não há avaliações
- Sistemas neuroendócrino e digestivoDocumento10 páginasSistemas neuroendócrino e digestivoFernando GuerraAinda não há avaliações
- FisiologiaDocumento1 páginaFisiologiadavibsouzaAinda não há avaliações
- Pâncreas - NEYDocumento3 páginasPâncreas - NEYChristian Zucolotto100% (1)
- ESTÔMAGODocumento20 páginasESTÔMAGOMichelle AraújoAinda não há avaliações
- O_pâncreas_é_um-WPS_OfficeDocumento12 páginasO_pâncreas_é_um-WPS_OfficeCalebe Geraldo DamiaoAinda não há avaliações
- Controle neuroendócrino da digestãoDocumento165 páginasControle neuroendócrino da digestãoMarcelo NévoaAinda não há avaliações
- Problema 8Documento12 páginasProblema 8Moises Popp de OliveiraAinda não há avaliações
- Sistema digestório emDocumento10 páginasSistema digestório emVeronicalg20Ainda não há avaliações
- Metabolismo dos macronutrientes no estado alimentadoDocumento6 páginasMetabolismo dos macronutrientes no estado alimentadoRaphael AntônioAinda não há avaliações
- Prova 3Documento24 páginasProva 3fehbondezanAinda não há avaliações
- Sistema Endócrino e SaciedadeDocumento77 páginasSistema Endócrino e SaciedadeTaphynes GuimarãesAinda não há avaliações
- Trab Renata BestDocumento4 páginasTrab Renata BestRaphael AlvesAinda não há avaliações
- Fisiologia Unidade4Documento21 páginasFisiologia Unidade4Juninho SilvaAinda não há avaliações
- Avaliação Laboratorial Hepatica e Pancreatica SANARDocumento33 páginasAvaliação Laboratorial Hepatica e Pancreatica SANAREderson ChavesAinda não há avaliações
- Bioquímica Insulina e Glicocorticoides Aula - 33Documento15 páginasBioquímica Insulina e Glicocorticoides Aula - 33Rodrigo Costa100% (1)
- CiênciasDocumento8 páginasCiênciasClarice AlmeidaAinda não há avaliações
- Aula 4 - LPF - Diabetes Gestacional - Teste de Tolerância Oral ÀDocumento11 páginasAula 4 - LPF - Diabetes Gestacional - Teste de Tolerância Oral Àgabriella.ferreira.inacioAinda não há avaliações
- Metabolismo e produção de calor: os processos de transformação de nutrientes em energiaDocumento6 páginasMetabolismo e produção de calor: os processos de transformação de nutrientes em energiaJulia Borsatto SchmitzAinda não há avaliações
- Problema 01 10Documento23 páginasProblema 01 10Karoline Silva ZampronhoAinda não há avaliações
- Golan 29Documento17 páginasGolan 29Manuela Rodrigues0% (1)
- Sistema DigestórioDocumento9 páginasSistema DigestórioenzoicafieroAinda não há avaliações
- Digestório 1Documento18 páginasDigestório 1Arthur MartinsAinda não há avaliações
- Tut VDocumento5 páginasTut VGabriel PiresAinda não há avaliações
- 80 FDDocumento7 páginas80 FDPedro CarlosAinda não há avaliações
- Diuréticos, rins e função renalDocumento9 páginasDiuréticos, rins e função renalFernand TananAinda não há avaliações
- 21 - Efeitos Metabólicos Da Insulina e Do Glucagon PDFDocumento3 páginas21 - Efeitos Metabólicos Da Insulina e Do Glucagon PDFLorenzo Salvestrini100% (1)
- THAIS RODRIGUES DE FARIA - FISIOLOGIA I - Estudo Dirigido M3 - Documentos GoogleDocumento3 páginasTHAIS RODRIGUES DE FARIA - FISIOLOGIA I - Estudo Dirigido M3 - Documentos GoogleThaís RodriguesAinda não há avaliações
- Revista Células PancreáticasDocumento34 páginasRevista Células Pancreáticasjessica.itaAinda não há avaliações
- Prova MetabolismoDocumento14 páginasProva MetabolismoiancacristalAinda não há avaliações
- Efeitos Metabólicos Da Insulina e Do GlucagonDocumento4 páginasEfeitos Metabólicos Da Insulina e Do Glucagonmarinadsena100% (1)
- Controle Neuroendocrino Da SaciedadeDocumento23 páginasControle Neuroendocrino Da Saciedadearthur hennys barbosaAinda não há avaliações
- Anatomia Do Sistema DigestórioDocumento3 páginasAnatomia Do Sistema DigestórioGptoAinda não há avaliações
- PâncreasDocumento3 páginasPâncreasGabriela Ferst De RéAinda não há avaliações
- EstomagoDocumento5 páginasEstomagoVictor PessoaAinda não há avaliações
- Fisiologia GastrointestinalDocumento4 páginasFisiologia GastrointestinalThayna CassulaAinda não há avaliações
- Controle Neuroend Crino Da SaciedadeDocumento22 páginasControle Neuroend Crino Da SaciedadelunarwopenAinda não há avaliações
- Ilhotas de Langerhans SlideDocumento2 páginasIlhotas de Langerhans Slidealineramona64Ainda não há avaliações
- Apostila DiabetesDocumento36 páginasApostila DiabetesCelso Murilo MartinsAinda não há avaliações
- OpenAccess Alvarez Leite 9788580391893 16Documento22 páginasOpenAccess Alvarez Leite 9788580391893 16Leonardo Azevedo100% (1)
- Controle neuroendócrino da saciedade e hormônios intestinaisDocumento22 páginasControle neuroendócrino da saciedade e hormônios intestinaisRicardo Wallace Das Chagas Lucas100% (1)
- Arquivo Seminario de FisiologiaDocumento6 páginasArquivo Seminario de FisiologiaDiovanamrs -Ainda não há avaliações
- Fisiologia Gastrointestinal - 4 Semestre - 2022Documento13 páginasFisiologia Gastrointestinal - 4 Semestre - 2022rafaelaAinda não há avaliações
- 1 Aula - Revisão Do Sistema DigestivoDocumento12 páginas1 Aula - Revisão Do Sistema DigestivoAlex SilvaAinda não há avaliações
- Células adiposas e suas funçõesDocumento23 páginasCélulas adiposas e suas funçõesLouis PeixotoAinda não há avaliações
- Anatomia e fisiologia do fígadoDocumento14 páginasAnatomia e fisiologia do fígadoAnna Belly '-'Ainda não há avaliações
- Prolactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoNo EverandProlactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoAinda não há avaliações
- Valvulopatias: Fisiologia, Anatomia e Sinais das Principais LesõesDocumento26 páginasValvulopatias: Fisiologia, Anatomia e Sinais das Principais LesõeselisaAinda não há avaliações
- Resumo Completo - Insuficiência CardíacaDocumento16 páginasResumo Completo - Insuficiência CardíacaelisaAinda não há avaliações
- TCE + HIC - Semana 4Documento21 páginasTCE + HIC - Semana 4elisaAinda não há avaliações
- Síndrome Coronariana Aguda e Infarto Agudo Do Miocárdio - Morfo1m1Documento22 páginasSíndrome Coronariana Aguda e Infarto Agudo Do Miocárdio - Morfo1m1elisaAinda não há avaliações
- Polígono de Willis e isquemia cerebralDocumento18 páginasPolígono de Willis e isquemia cerebralelisaAinda não há avaliações
- Asma e Pneumonia - Morfo 2Documento27 páginasAsma e Pneumonia - Morfo 2elisaAinda não há avaliações
- Cap 3Documento33 páginasCap 3Juliana OliveiraAinda não há avaliações
- Metabolismo Celular 1 Ano Aula 01Documento22 páginasMetabolismo Celular 1 Ano Aula 01Nulciene FreitasAinda não há avaliações
- Questionário de BioquímicaDocumento13 páginasQuestionário de BioquímicajosielquiAinda não há avaliações
- Aula 02 - Metabolismo de CarboidratosDocumento36 páginasAula 02 - Metabolismo de CarboidratosCilene LatorracaAinda não há avaliações
- Metabolismo dos hidratos de carbono: glicogénioDocumento28 páginasMetabolismo dos hidratos de carbono: glicogénioDiogoFevereiroAinda não há avaliações
- Fermentação de açúcares por PasteurDocumento3 páginasFermentação de açúcares por Pasteursonia silvaAinda não há avaliações
- Água e suas funções no organismoDocumento8 páginasÁgua e suas funções no organismovalentinAinda não há avaliações
- Bioquímica Área IIDocumento35 páginasBioquímica Área IINilza UFRGSAinda não há avaliações
- Respiração celular: glicólise e substratosDocumento21 páginasRespiração celular: glicólise e substratosDaniela ChavesAinda não há avaliações
- PAES 2015 - Questões de BiologiaDocumento27 páginasPAES 2015 - Questões de BiologiaCaio RafaelAinda não há avaliações
- Bioquímica e MetabolismoDocumento26 páginasBioquímica e MetabolismoJervàcio ChambalAinda não há avaliações
- Bioenergética, sistemas energéticos e substratos no exercícioDocumento6 páginasBioenergética, sistemas energéticos e substratos no exercícioGreicy Peretti PoffoAinda não há avaliações
- Caso Clínico - Glicogenose - 2023Documento9 páginasCaso Clínico - Glicogenose - 2023vanessa.vancoAinda não há avaliações
- Apostila completa sobre sistemas energéticos, recuperação muscular e treino idealDocumento58 páginasApostila completa sobre sistemas energéticos, recuperação muscular e treino idealajaquilanteAinda não há avaliações
- Aula Metabolismo Microbiano Parte BDocumento46 páginasAula Metabolismo Microbiano Parte BMilton Luiz da Paz LimaAinda não há avaliações
- Mecanismos Geração EnergiaDocumento13 páginasMecanismos Geração EnergiaRicardo GuraAinda não há avaliações
- Questionário - Questões Marks - 2 ProvaDocumento8 páginasQuestionário - Questões Marks - 2 ProvaJohanaAinda não há avaliações
- BioquímicaDocumento4 páginasBioquímicaAliny Antunes Lobo LaddAinda não há avaliações
- BIOENERGÉTICA - CópiaDocumento37 páginasBIOENERGÉTICA - CópiaMaria Luíza AguiarAinda não há avaliações
- Livro de Fisicologia 2015Documento140 páginasLivro de Fisicologia 2015Maria Rosario Fatima SousaAinda não há avaliações
- Relatório de Aula Prática FermentaçãoDocumento5 páginasRelatório de Aula Prática FermentaçãoWend MairaAinda não há avaliações