Você também pode gostar
- Farmacodinâmica e interação fármaco-receptorDocumento37 páginasFarmacodinâmica e interação fármaco-receptorVictor MoreiraAinda não há avaliações
- Farmacodinâmica-Interação fármaco-receptorDocumento11 páginasFarmacodinâmica-Interação fármaco-receptorBruna GomesAinda não há avaliações
- Questoes Farmacodinamica GabaritoDocumento3 páginasQuestoes Farmacodinamica GabaritoBruna Lucas Dal MolinAinda não há avaliações
- Introdução à FarmacologiaDocumento137 páginasIntrodução à Farmacologiacris100% (2)
- Resumo Farmacodinâmica e FarmacocinéticaDocumento16 páginasResumo Farmacodinâmica e FarmacocinéticaMaiara Perussolo100% (1)
- FARMACOLOGIADocumento6 páginasFARMACOLOGIABruna KelletAinda não há avaliações
- Atividade FarmacodinamicaDocumento3 páginasAtividade FarmacodinamicaVitoria AlmeidaAinda não há avaliações
- 1552949292-Farmacologia Principios BasicosDocumento24 páginas1552949292-Farmacologia Principios BasicosBrunaLuízaAinda não há avaliações
- Farmacocinêt e farmacodinâmicaDocumento15 páginasFarmacocinêt e farmacodinâmicaAna GrozinskiAinda não há avaliações
- Guia Completo das Terapias Alternativas: Métodos terapêuticos naturais que proporcionam saúde integralNo EverandGuia Completo das Terapias Alternativas: Métodos terapêuticos naturais que proporcionam saúde integralNota: 4 de 5 estrelas4/5 (15)
- Farmacodinâmica Dos MedicamentosDocumento11 páginasFarmacodinâmica Dos MedicamentosCélïö100% (1)
- Farmacologia e Mecanismos de AçãoDocumento31 páginasFarmacologia e Mecanismos de AçãoBruno DiasAinda não há avaliações
- Farmacodinâmica SagahDocumento24 páginasFarmacodinâmica Sagahjayh0p394Ainda não há avaliações
- FarmacocinéticaDocumento10 páginasFarmacocinéticaLília BoellAinda não há avaliações
- 3 - FarmacodinâmicaDocumento13 páginas3 - FarmacodinâmicaGabriele BassoAinda não há avaliações
- FarmacologiaDocumento19 páginasFarmacologiaLuis BigheadAinda não há avaliações
- 4 - FarmacodinâmicaDocumento30 páginas4 - FarmacodinâmicaWendell OliveiraAinda não há avaliações
- Curso Psicofarmacologia EADDocumento17 páginasCurso Psicofarmacologia EADGisleneAlvesAinda não há avaliações
- Mecanismo de Ação Dos FármacosDocumento7 páginasMecanismo de Ação Dos FármacosLarissa GabrielleAinda não há avaliações
- Farmacodinâmica: Estudo dos Efeitos dos FármacosDocumento20 páginasFarmacodinâmica: Estudo dos Efeitos dos FármacosSilvania MartinsAinda não há avaliações
- Aula 03 - Farmacodinâmica (1) .PPTX Pit 01-22Documento48 páginasAula 03 - Farmacodinâmica (1) .PPTX Pit 01-22Hdms S100% (2)
- Apostila de FarmacologiaDocumento45 páginasApostila de FarmacologiaJaqueline BarbosaAinda não há avaliações
- Apostila Farmaco I UNIRIODocumento45 páginasApostila Farmaco I UNIRIOesdrassantanaAinda não há avaliações
- Farmacologia E-book: Introdução à FarmacologiaDocumento62 páginasFarmacologia E-book: Introdução à Farmacologiavalentin100% (4)
- FarmacodinâmicaDocumento6 páginasFarmacodinâmicaGabriela Maria Bastos dos SantosAinda não há avaliações
- Manual Recke We GDocumento283 páginasManual Recke We Gsoraia100% (5)
- Noções de farmacologiaDocumento96 páginasNoções de farmacologiaVitória MoreiraAinda não há avaliações
- Entendendo a farmacologia básica e psicofarmacologia das bulasDocumento17 páginasEntendendo a farmacologia básica e psicofarmacologia das bulasLia MartinsAinda não há avaliações
- Apostila ProntaDocumento38 páginasApostila ProntaClaudileide CazavechiaAinda não há avaliações
- Farmacologia I.Documento76 páginasFarmacologia I.220217240% (1)
- Roteiro II de FarmacologiaDocumento4 páginasRoteiro II de FarmacologiaPEDRO HENRIQUE FERREIRA GUIMARÃESAinda não há avaliações
- SafariDocumento7 páginasSafariMurilo Spricigo GeremiasAinda não há avaliações
- AULA Farmacocinetica e FarmacodinamicaDocumento17 páginasAULA Farmacocinetica e FarmacodinamicaJosiane Gama0% (1)
- Questões de QuimicaDocumento3 páginasQuestões de QuimicaAndré Silva MoraisAinda não há avaliações
- Cap 1 ApostilaDocumento13 páginasCap 1 Apostilaandressa Leal -CCB- ParáAinda não há avaliações
- Farmacologia - As vertentes e definiçõesDocumento58 páginasFarmacologia - As vertentes e definiçõesGivanildo BenicioAinda não há avaliações
- Interacao farmaco-receptor: agonistas, antagonistas e efeitosDocumento4 páginasInteracao farmaco-receptor: agonistas, antagonistas e efeitosÉrika NascimentoAinda não há avaliações
- FARMACODINÂMICADocumento34 páginasFARMACODINÂMICAviber2525Ainda não há avaliações
- Introdução À FarmacologiaDocumento28 páginasIntrodução À FarmacologiaWêndeo CostaAinda não há avaliações
- Apresentação 3Documento12 páginasApresentação 3Pamela GrangeiroAinda não há avaliações
- Farmacologia: Mecanismo de Ação e Classificação dos FármacosDocumento5 páginasFarmacologia: Mecanismo de Ação e Classificação dos Fármacosmailson eleuterioAinda não há avaliações
- Nocões de Farmacologia Aula 1Documento28 páginasNocões de Farmacologia Aula 1Fernando FelippeAinda não há avaliações
- Apostila 01 Introducao A FarmacologiaDocumento6 páginasApostila 01 Introducao A FarmacologiaWilma de JesusAinda não há avaliações
- Introdução à Farmacologia I - Receptores e Interações Drogas-ReceptorDocumento41 páginasIntrodução à Farmacologia I - Receptores e Interações Drogas-ReceptorRafaSoaresAinda não há avaliações
- Aula 01 FarmacologiaDocumento44 páginasAula 01 FarmacologiaÍtala Barbosa MachadoAinda não há avaliações
- 1 4902701405992649100Documento1.227 páginas1 4902701405992649100klausAinda não há avaliações
- Farmacologia Aplicada A OdontologiaDocumento91 páginasFarmacologia Aplicada A OdontologiaJúlio LaraAinda não há avaliações
- Aula 2 FarmacodinâmicaDocumento48 páginasAula 2 FarmacodinâmicaGisele FernandesAinda não há avaliações
- Farmacologia Traduzida SebentaDocumento200 páginasFarmacologia Traduzida SebentaSofia VilarAinda não há avaliações
- 1 Aula Teórica de Farmacodinamia (3)Documento23 páginas1 Aula Teórica de Farmacodinamia (3)Giovana BragaAinda não há avaliações
- TEMA 1 - Parte III - Fundamentos Da Farmacologia - FARMACODINÂMICADocumento33 páginasTEMA 1 - Parte III - Fundamentos Da Farmacologia - FARMACODINÂMICAdiasluiza863Ainda não há avaliações
- Aula 1 FarmacocinéticaDocumento67 páginasAula 1 FarmacocinéticaGisele FernandesAinda não há avaliações
- 1 - Introdução À HomotoxicologiaDocumento60 páginas1 - Introdução À Homotoxicologiarui_terrinha3041100% (2)
- 01 - Introdução À FarmacologiaDocumento6 páginas01 - Introdução À FarmacologiaThiana RamosAinda não há avaliações
- Farmacologia: Farmacodinâmica e FarmacocinéticaDocumento20 páginasFarmacologia: Farmacodinâmica e FarmacocinéticaGUILHERME SILVA MARTINS SANTOSAinda não há avaliações
- -farmacologia aula 2Documento60 páginas-farmacologia aula 2layanasasilvaAinda não há avaliações
- Introdução aos Agonistas e AntagonistasDocumento3 páginasIntrodução aos Agonistas e AntagonistasVictória AkkariAinda não há avaliações
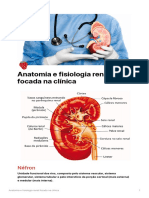
- Anatomia e Fisiologia Renal Focada Na ClnicaDocumento7 páginasAnatomia e Fisiologia Renal Focada Na ClnicaEnzo RodriguesAinda não há avaliações
- Artrite Reumatoide: Epidemiologia, Clínica, Diagnóstico e TratamentoDocumento7 páginasArtrite Reumatoide: Epidemiologia, Clínica, Diagnóstico e TratamentoEnzo RodriguesAinda não há avaliações
- Transtornos AnsiososDocumento5 páginasTranstornos AnsiososEnzo RodriguesAinda não há avaliações
- Transtornos de HumorDocumento7 páginasTranstornos de HumorEnzo RodriguesAinda não há avaliações
- TRUSPA: Transtornos Relacionados ao Uso de Substâncias PsicoativasDocumento4 páginasTRUSPA: Transtornos Relacionados ao Uso de Substâncias PsicoativasEnzo RodriguesAinda não há avaliações
- Psicoses: sinais, causas e tratamentoDocumento7 páginasPsicoses: sinais, causas e tratamentoEnzo RodriguesAinda não há avaliações
- HIPEREMIA, CONGESTÃO E EDEMA: ALTERAÇÕES HEMODINÂMICASDocumento5 páginasHIPEREMIA, CONGESTÃO E EDEMA: ALTERAÇÕES HEMODINÂMICASEnzo RodriguesAinda não há avaliações
- ReparoDocumento4 páginasReparoEnzo RodriguesAinda não há avaliações
- Adaptações CelularesDocumento4 páginasAdaptações CelularesEnzo RodriguesAinda não há avaliações
- Lesões celulares reversíveis e degeneração gordurosaDocumento4 páginasLesões celulares reversíveis e degeneração gordurosaEnzo RodriguesAinda não há avaliações
- Sinalização intercelular e transdução de sinalDocumento27 páginasSinalização intercelular e transdução de sinalCarina Barbosa100% (1)
- Sinalização Molecular e Receptores CelularesDocumento736 páginasSinalização Molecular e Receptores CelularesRicardo MacedoAinda não há avaliações
- Sinalização CelularDocumento19 páginasSinalização CelularFlavio Tino dos JoaoAinda não há avaliações
- Cederj - Bio Cel I - Vol 2Documento133 páginasCederj - Bio Cel I - Vol 2Camille PradoAinda não há avaliações
- Farmacologia ReceptoresDocumento46 páginasFarmacologia ReceptorescamilleAinda não há avaliações
- Curso de Farmacologia Básica com Fisiologia DetalhadaDocumento48 páginasCurso de Farmacologia Básica com Fisiologia Detalhadaandre almeidaAinda não há avaliações
- Sinal CelularDocumento22 páginasSinal CelularBruno CordeiroAinda não há avaliações
- 1.2 Mecanismo de Transdução HormonalDocumento3 páginas1.2 Mecanismo de Transdução HormonalJaqueline SantosAinda não há avaliações
- Ligação iônica e covalente, propriedades da água e lei de Lambert-BeerDocumento4 páginasLigação iônica e covalente, propriedades da água e lei de Lambert-BeerVictoria MendesAinda não há avaliações
- Banco de Questes de Farmacologia PDFDocumento26 páginasBanco de Questes de Farmacologia PDFCristine Dantas Lima83% (6)
- ED FarmacodinâmicaDocumento2 páginasED FarmacodinâmicaFelipe Fonseca SilvaAinda não há avaliações
- Cap 74 - Introdução A EndocrinologiaDocumento7 páginasCap 74 - Introdução A Endocrinologiaisabella dacroceAinda não há avaliações
- Estudo Dirigido Sinalização e Diferenciação Celular PDFDocumento1 páginaEstudo Dirigido Sinalização e Diferenciação Celular PDFAvguzto Plinio Souza100% (1)
- Farmacodinâmica II - Mecanismo de Ação de DrogasDocumento14 páginasFarmacodinâmica II - Mecanismo de Ação de DrogasRaul Carpi SantosAinda não há avaliações
- 2 Aula FarmadinamicaDocumento48 páginas2 Aula FarmadinamicaEduardo CostaAinda não há avaliações
- Princípios fundamentais de farmacologia: Interações fármaco-receptorDocumento16 páginasPrincípios fundamentais de farmacologia: Interações fármaco-receptorDaniel Sant AnaAinda não há avaliações
- Resumo Bioquímica Mecanismos de Ação HormonalDocumento7 páginasResumo Bioquímica Mecanismos de Ação HormonalMariane KolandjianAinda não há avaliações
- UFAC 2014dDocumento16 páginasUFAC 2014dLUCES Liga Universitaria de Cirugías y EstéticaAinda não há avaliações
- Vias de Administração e FarmacodinamicaDocumento26 páginasVias de Administração e FarmacodinamicaZoeAinda não há avaliações
- Receptores de Membrana e Acoplados À Proteina GDocumento8 páginasReceptores de Membrana e Acoplados À Proteina GMax Nunes100% (2)
- Mecanismos de ação das drogasDocumento5 páginasMecanismos de ação das drogasWeslany SyllvaAinda não há avaliações
- AEPs Geral M8Documento64 páginasAEPs Geral M8Carolina ChongAinda não há avaliações
- Aula 6 - Transdução de Sinais BiológicosDocumento12 páginasAula 6 - Transdução de Sinais BiológicosJailton MagalhãesAinda não há avaliações
- Comunicação celular: receptores e sinalizaçãoDocumento6 páginasComunicação celular: receptores e sinalizaçãoEnzoAinda não há avaliações
- Psicofarmacologia: conceitos e históriaDocumento162 páginasPsicofarmacologia: conceitos e históriaQuéssia RochaAinda não há avaliações
- Mapa de Sinalização CelularDocumento3 páginasMapa de Sinalização CelularLana OtoniAinda não há avaliações
- Analgésicos opióides: mecanismos, tipos e usosDocumento33 páginasAnalgésicos opióides: mecanismos, tipos e usosDenise Terenzi100% (1)
- Transtorno BipolarDocumento30 páginasTranstorno BipolardesireepaysageAinda não há avaliações
- 6 - Princípios Da Ação Dos Fármacos 2023Documento31 páginas6 - Princípios Da Ação Dos Fármacos 2023Rayssa SilvaAinda não há avaliações
- Exercicios Terceiro ModuloDocumento3 páginasExercicios Terceiro ModuloGabriela AndradeAinda não há avaliações